
Bill Gates: Milde Omicron Variante wird akute Phase der Pandemie 2022 beenden
Impfzwang unnötig
Impfzwang nicht nötig: Omicron ist leichte Variante und fast sicher - so Fauci
Durchseuchung mit milder Omicron Variante sei wünschenswert
Der oberste amerikanische Impf-Papst Anthony Fauci bezeichnet die neue Omocron-Variante des Coronavirus als fast sicher und als relativ mild.
Eine Durchseuchung des Volkes mit dieser milden Variante sei sogar wünschenswert. Omicron könnte im Vergleich zu anderen Varianten sogar zu einer milderen Krankheit führen.
Nach diesen Aussagen von Fauci waren Weltbörsen wieder stark gestiegen, nachdem sie nach Auftauchen der Omicron Variante zunächst stark gefallen waren.
Bereits am Anfang hatte man mitbekommen, dass diese Variante wie in Botswana eigentlich nur Geimpfte betrifft.
Andererseits haben englische Daten der Gesundheitsbehörden klar gemacht, dass 80 % der Coronatoten voll geimpfte Menschen sind.
Zudem zeigen weltweite Untersuchungen, dass eine Massendurchimpfung von über 30 % der Menschen kontraproduktiv ist und das Virus zu Fluchtmutationen treibt, die dann wieder viel gefährlicher sein könnten als bisherige Varianten. Zudem ist die Zahl der Corona Erkrankungen laut amerikanischen Studien je 1 Mio Einwohner umso höher , desto größer die Impfquote ist.
Auch die allgemeine Todesrate ist bei Geimpften laut Harvard Studie 6 mal höher als bei Ungeimpften. In Deutschland kommt es wegen der global betrachtet viel zu hohen Impfquote von über 60 % zu einer Übersterblichkeit in allen 16 Bundesländern.
Desweiteren wirkt der herkömmliche Impfstoff überhaupt nicht gegen die Omikron-Variante, so dass alle 70 % Geimpften und Geboosterten erneut geimpft werden müssten. Die Wirkung soll um mindestens 37 fache Reduktion sinken.
So gesehen ist ein allgemeiner Impfzwang nicht nur nicht notwendig sondern schädlich.
Impfzwang nicht nötig: Omicron ist leichte Variante und fast sicher - so Fauci
Durchseuchung mit milder Omicron Variante sei wünschenswert
Der oberste amerikanische Impf-Papst Anthony Fauci bezeichnet die neue Omocron-Variante des Coronavirus als fast sicher und als relativ mild.
Eine Durchseuchung des Volkes mit dieser milden Variante sei sogar wünschenswert. Omicron könnte im Vergleich zu anderen Varianten sogar zu einer milderen Krankheit führen.
Nach diesen Aussagen von Fauci waren Weltbörsen wieder stark gestiegen, nachdem sie nach Auftauchen der Omicron Variante zunächst stark gefallen waren.
Bereits am Anfang hatte man mitbekommen, dass diese Variante wie in Botswana eigentlich nur Geimpfte betrifft.
Andererseits haben englische Daten der Gesundheitsbehörden klar gemacht, dass 80 % der Coronatoten voll geimpfte Menschen sind.
Zudem zeigen weltweite Untersuchungen, dass eine Massendurchimpfung von über 30 % der Menschen kontraproduktiv ist und das Virus zu Fluchtmutationen treibt, die dann wieder viel gefährlicher sein könnten als bisherige Varianten. Zudem ist die Zahl der Corona Erkrankungen laut amerikanischen Studien je 1 Mio Einwohner umso höher , desto größer die Impfquote ist.
Auch die allgemeine Todesrate ist bei Geimpften laut Harvard Studie 6 mal höher als bei Ungeimpften. In Deutschland kommt es wegen der global betrachtet viel zu hohen Impfquote von über 60 % zu einer Übersterblichkeit in allen 16 Bundesländern.
Desweiteren wirkt der herkömmliche Impfstoff überhaupt nicht gegen die Omikron-Variante, so dass alle 70 % Geimpften und Geboosterten erneut geimpft werden müssten. Die Wirkung soll um mindestens 37 fache Reduktion sinken.
So gesehen ist ein allgemeiner Impfzwang nicht nur nicht notwendig sondern schädlich.
Angela Merkel zieht in Margot Honeckers Büro
Zurück zu den Wurzeln
Ob Ex Kanzlerin Angela Merkel nun Stasi IM in DDR Zeiten war, verrät sie trotz der vielen Indizien vorerst nicht.
Aber sie hatte sich zum Abschied schon den DDR Schlager von Nina Hagen " Du hast den Farbfilm vergessen Michael" gewünscht.
Nur wenige Zeitgenossen wissen, dass sie in Russland zeitweise studierte und dort ihren ehemaligen Lebensgefährten Michael Schindhelm traf, der trotz jahrelanger Leugnung dann Jahre nach der Wende doch als Stasi IM des Mielke-Ministeriums der DDR enttarnt wurde und einen Exil-Job als Theaterintendant in Basel erhalten und sozusagen weggelobt wurde - obwohl er in der DDR nur sehr marginal künstlerisch allerdings mit Verweis im Roman auf Angela Merkel in Erscheinung trat.
Auch Angela Merkel tauchte in brisanten Stasiakten wie der Akte Robert Havemann mit unbekannter Funktion auf. Havemann war jahrelang in Isolationshaft in Dauerbeobachtung durch ganze Stasi-Jugendbrigaden .
Angela Merkel war als stellvertretende FDJ Vorsitzende neben Professor Hans Jörg Osten, der Erster Sekretär an der Akademie der Wissenschaften der DDR war, praktisch seine rechte Hand. Dort lehrte er auch Marxismus -Leninismus und er besetzte mit Angela Merkel zusammen Häuser in Ost-Berlin . Aber vor allem stellte sich später heraus, dass Hans Jörg Osten nicht nur ein Stasi IM war sondern wahrscheinlich der wichtigste Stasi Agent der DDR, der den Ehrentitel IM Einstein erhalten hatte.
Jetzt setzt Angela Merkel jedenfalls noch eine drauf und rühmt sich damit, die Honeckers "zu beerben".
Angela Merkel zieht in "Margot Honeckers Büro"

Angela Merkels Feinde spotteten schon immer, sie sei "Erich Honeckers letzte Rache", und noch zu Beginn ihrer Kanzlerschaft legte die Satiresendung "Mitternachtsspitzen" Wladimir Putin die Aussage in den Mund, dies sei die ultimative Geheimwaffe des Ostblocks, "die den Westen für immer fertig machen wird."
Doch das hier ist nun weder Witz noch Spötterei: Niemand anderes als die scheidende Bundeskanzlerin selbst prahlte bei ihrem letzten Treffen mit der CDU-Bundestagsfraktion am 7. Dezember, sie sei ja nicht aus der Welt, sondern ziehe in "Margot Honeckers Büro, wo auch Helmut Kohls Büro als Altkanzler gewesen ist."
Dies berichten heute übereinstimmend mehrere renommierte deutsche Presseerzeugnisse unter Berufung auf Bundestagsabgeordnete der CDU.
Das Bürohaus Ecke Wilhelmstraße / Unter den Linden in Berlin-Mitte wurde in den 1960er Jahren für das Ministerium für Volksbildung der DDR errichtet. Ausgestattet mit exklusivem Blick auf das Brandenburger Tor und das Stückchen Westberlins, das unmittelbar dahinter lag, residierte Margot Honecker hier von 1963 bis zum politischen Sturz ihres Ehemannes im Jahr 1989.

Nach dem Beitritt der DDR zur Bundesrepublik Deutschland im Oktober 1990 war das Gebäude zunächst Sondervermögen, und kam nach dem Umzug des Bundestages nach Berlin in die Verfügung der Bundestagsverwaltung. Der Blick auf das Brandenburger Tor ist nun allerdings durch die Neubauten der Britischen Botschaft und des Hotels Adlon verstellt, und kein Abgeordneter reißt sich mehr um ein Büro mit DDR-Zuschnitt. Dennoch ein würdiger Alterssitz für Deutschlands erste Alt-Bundeskanzlerin.
666 Fälle von Herzerkrankungen bei 12- bis 17-Jährigen nach COVID-Impfungen –
2.809 tote Babys in VAERS nach COVID-Impfungen, wobei neue Dokumente beweisen, dass Pfizer, die FDA und die CDC wussten, dass die Schüsse für schwangere Frauen nicht sicher sind
Weniger als 2 Fälle pro Jahr nach allen Impfstoffen in den letzten 30+ Jahren
Die COVID-19-Spritzen verursachen Herzkrankheiten, hauptsächlich Myokarditis und Perikarditis, die die Gesundheit unserer jungen Leute zerstört.
Dies ist eine Tatsache, die nicht mehr bestritten wird, denn selbst die CDC gibt dies zu, wie ihr jüngster Bericht feststellt :
Mit Stand vom 24. November 2021 hat VAERS 1.949 Berichte über Myokarditis oder Perikarditis bei Personen im Alter von 30 Jahren und jünger erhalten, die einen COVID-19-Impfstoff erhalten haben. Die meisten Fälle wurden nach der mRNA-COVID-19-Impfung (Pfizer-BioNTech oder Moderna) gemeldet, insbesondere bei männlichen Jugendlichen und jungen Erwachsenen. ( Quelle .)
Die einzigen strittigen Punkte sind, 1, ob diese Fälle „selten“ sind oder nicht, und 2, ob der Nutzen der COVID-19-Massenimpfung junger Menschen das Risiko für Herzerkrankungen überwiegt.
Und in diesen beiden Punkten belügt die CDC die Öffentlichkeit, wie ich in diesem Artikel schlüssig beweisen werde.
Der zweite Punkt ist eigentlich sehr leicht zu entlarven, indem man sich einfach öffentlich zugängliche Statistiken über COVID-19-Todesfälle für diese Altersgruppe ansieht.
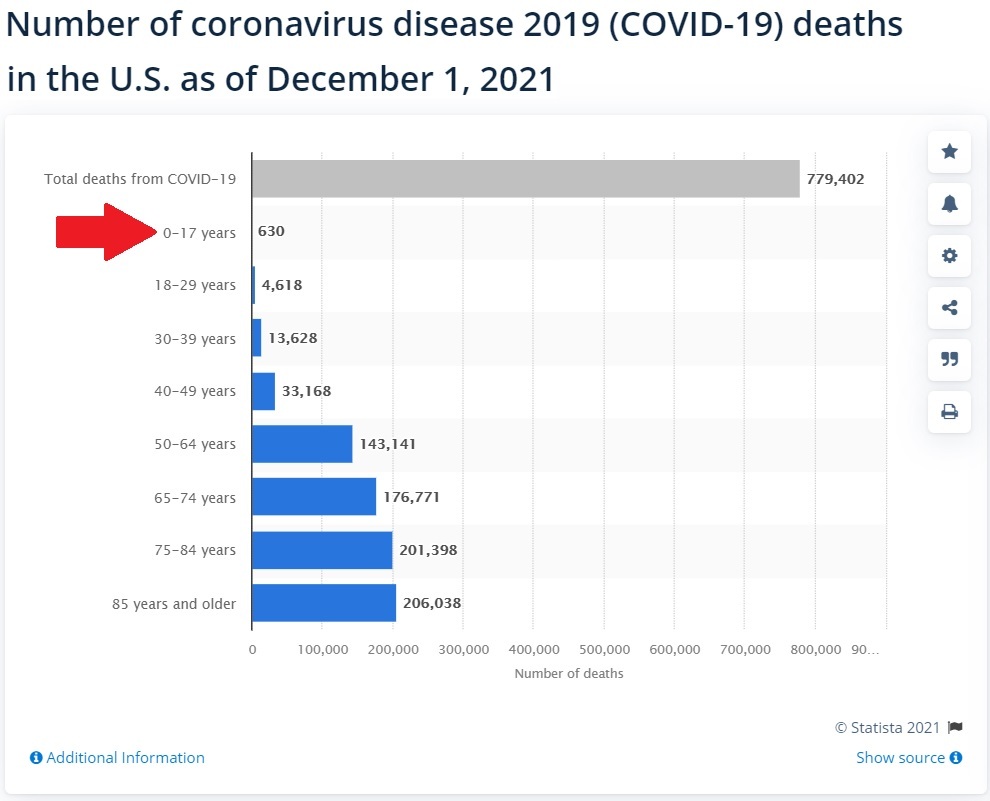
Quelle .
Am 1. Dezember 2021 waren von 779.402 mutmaßlichen COVID-19-Todesfällen, die jetzt fast 2 Jahre abdecken, nur 630 davon unter 17 Jahre alt.
Und selbst diese 630 angeblichen Todesfälle in dieser Altersgruppe werden nicht unbedingt durch COVID-19 verursacht. Es bedeutet nur, dass sie bei ihrem Tod positiv auf COVID-19 getestet wurden.
Es hat also keinen Vorteil, Kinder unter 17 Jahren gegen COVID-19 zu impfen, wenn sie statistisch eine nahezu nullprozentige Wahrscheinlichkeit haben, an COVID-19 zu sterben, wenn bekannt ist, dass diese Impfungen Herzkrankheiten verursachen.
In Bezug auf die Behauptung der CDC, dass Fälle von Herzerkrankungen, die durch COVID-19-Spritzen verursacht werden, „selten“ sind, sagen die Fakten etwas anderes.
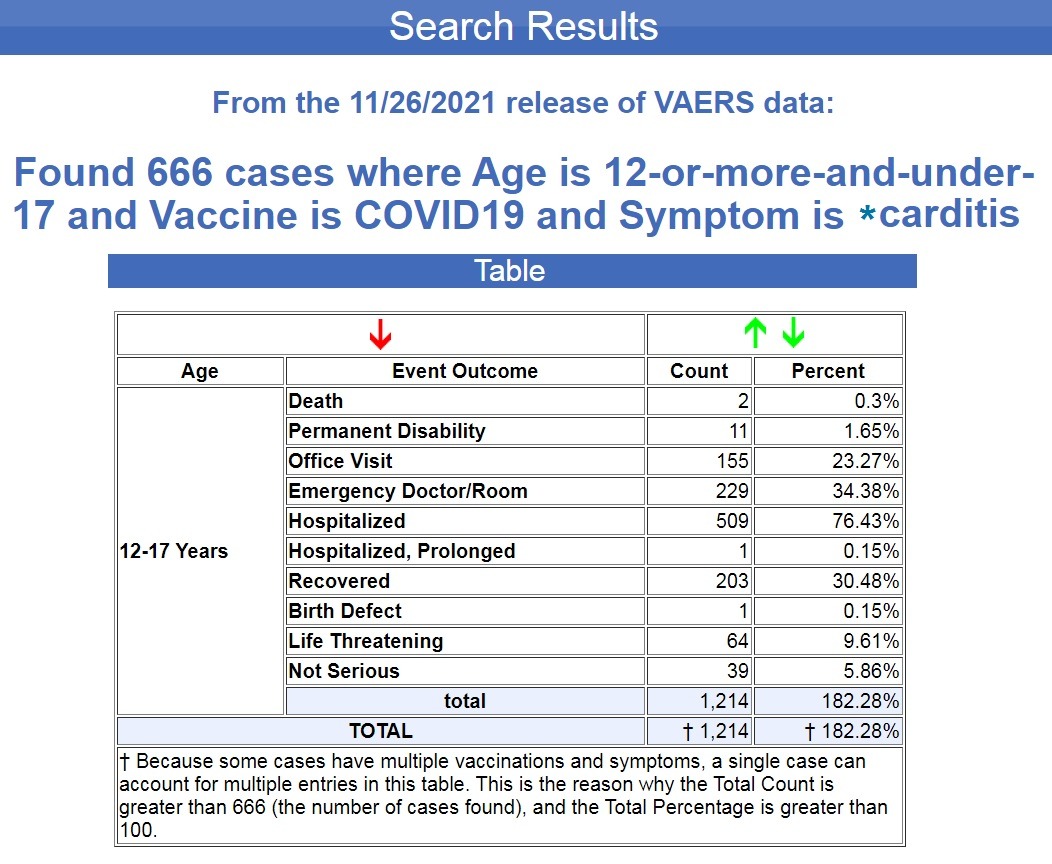
Ich habe in VAERS, dem Meldesystem für Impfstoffe gegen unerwünschte Ereignisse der US-Regierung , nach allen Fällen von „Karditis“ nach COVID-19-Spritzen für diese Altersgruppe gesucht, und seit der Veröffentlichung der VAERS-Daten vom 26.11.2021 wurde ein Ergebnis zurückgegeben von 666 Fällen . ( Quelle .)
Als nächstes führte ich genau dieselbe Suche nach derselben Altersgruppe durch, ausgenommen COVID-19-Impfungen und einschließlich aller anderen Impfstoffe, die in den letzten 30+ Jahren in VAERS aufgeführt waren, und ergab einen Wert von 40 Fällen, weniger als 2 Fälle pro Jahr. ( Quelle .)
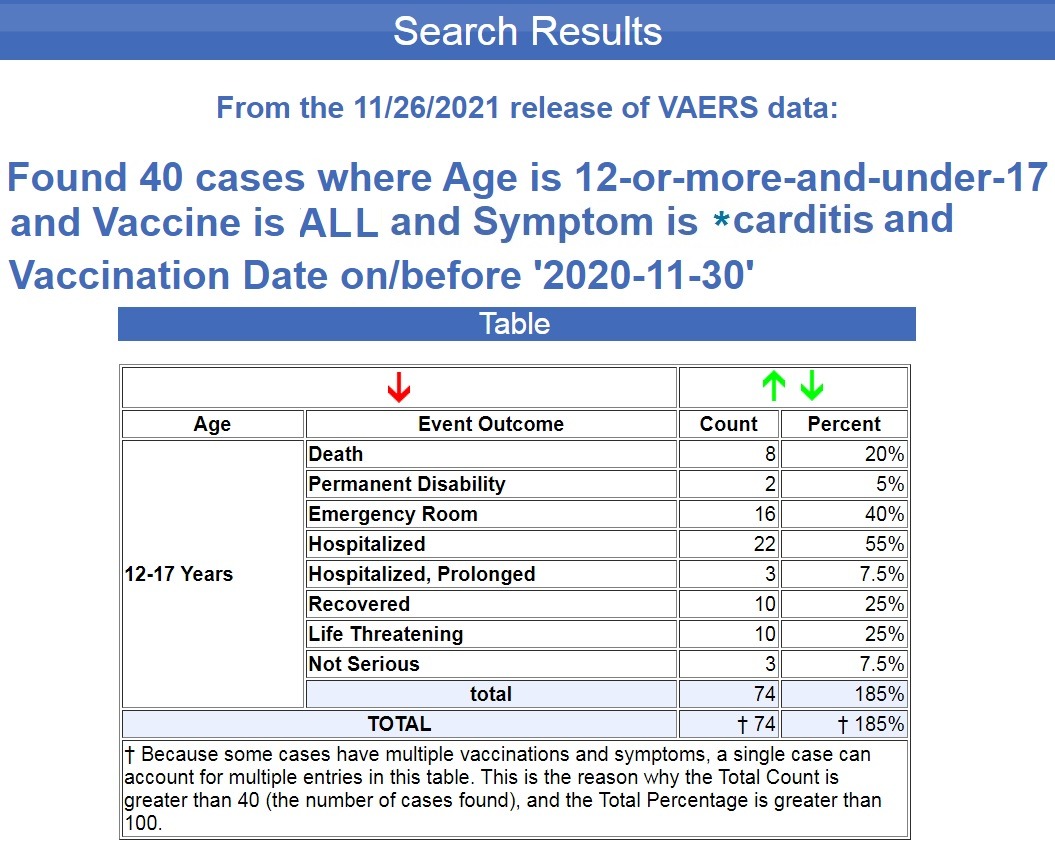
Denken Sie daran, dass die 666 Fälle in dieser Altersgruppe nach COVID-19-Impfungen für 12- bis 15-Jährige in den USA nur 6 Monate dauern, da die Pfizer-Impfung erst im Mai für diese Altersgruppe eine Notfallgenehmigung erhielt dieses Jahr (2021).
Kinder im Alter zwischen 5 und 11 Jahren erhielten letzten Monat, November 2021, gerade eine Notfallgenehmigung, aber es wurden bereits 2 Fälle von Herzerkrankungen in dieser Altersgruppe gemeldet, ein 6-jähriger und ein 8-jähriger, beide Jungen. ( Quelle .)
Nach wessen Definition sind diese Fälle von Herzerkrankungen bei diesen Kindern „selten“?

Wir haben vor ein paar Wochen berichtet, dass die Zeitschrift der American Heart Association, Circulation , eine Zusammenfassung des Kardiologen Steven R. Gundry veröffentlicht hat, in der behauptet wird, dass das Erhalten von COVID-19-Spritzen das Risiko für Herzinfarkte „dramatisch erhöht“.
Der britische Arzt Vernon Coleman sagte in Bezug auf die Studie: Endlich! Medizinischer Beweis für Covid Jab ist „Mord“ . Siehe: American Heart Association Journal veröffentlicht Daten, wonach britische Ärzte behaupten, dass sie „Beweise“ sind, dass COVID-19-Impfstoffe „Mord“ sind
Dr. Aseem Malhotra, ein weiterer Kardiologe aus Großbritannien, bestätigte die Ergebnisse der AHA-Studie und teilte mit, dass britische Behörden auf dem Gebiet der Kardiologie ihm bestätigten, dass dies geschieht, dass die COVID-19-Spritzen zu vermehrten Herzinfarkten führen, aber sie haben Angst, an die Öffentlichkeit zu gehen, weil sie ihre Forschungsgelder von den Arzneimittelherstellern verlieren.
Er forderte ein sofortiges Ende der Impfstoffmandate. Sehen Sie sich das Interview auf unserem Bitchute-Kanal an .
Ben Madgen: Ehemaliger Profi-Basketballspieler in Australien. Bildquelle .
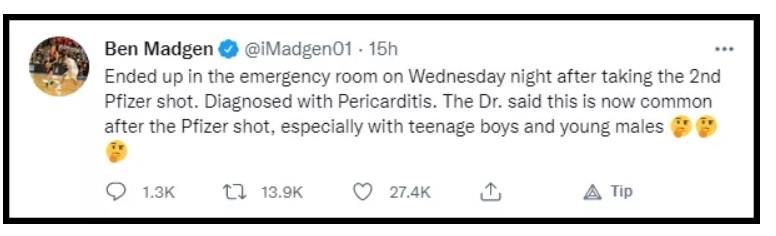
Die COVID-Welt berichtete kürzlich, dass bei dem ehemaligen australischen Profi-Basketballspieler Ben Madgen kurz nach Erhalt seiner zweiten Dosis des Pfizer-COVID-19-Impfstoffs eine Perikarditis (Entzündung des das Herz umgebenden Sacks) diagnostiziert wurde.
Die australischen Ärzte, die ihn behandelten, stimmen offenbar nicht mit der CDC überein, dass dies eine „seltene“ Nebenwirkung ist, da sie ihm sagten, dass Perikarditis jetzt eine häufige Nebenwirkung der Pfizer-Spritze bei jungen Männern ist.
Es gibt mittlerweile so viele Berichte über Sportler und andere junge Männer, die seit Beginn der COVID-19-Spritzen an Herzinfarkten leiden, dass die medizinischen Behörden versuchen mussten, eine andere Erklärung zu finden, um die Aufmerksamkeit von den tödlichen COVID-19-Spritzen abzulenken.
Also erfanden sie eine neue Krankheit: „Post-Pandemie-Belastungsstörung“.
Bis zu 300.000 Menschen in Großbritannien sind aufgrund einer postpandemischen Belastungsstörung (PPSD) mit Herzerkrankungen konfrontiert, warnten zwei Londoner Ärzte.
Dies könnte aufgrund der Auswirkungen von PPSD zu einem landesweiten Anstieg der kardiovaskulären Fälle um 4,5 Prozent führen, wobei die Altersgruppe zwischen 30 und 45 Jahren am stärksten gefährdet ist.
Mark Rayner, ein ehemaliger leitender psychologischer NHS-Therapeut und Gründer von EASE Wellbeing CIC, sagte, dass aufgrund von Stress und Angstzuständen, die durch die Auswirkungen von Covid-19 verursacht werden, bereits bis zu drei Millionen Menschen in Großbritannien an PPSD leiden. ( Quelle .)
Das kannst du dir nicht ausdenken!
Wenn Erwachsene dumm genug sein wollen, um diesen Unsinn zu glauben und eine COVID-19-Spritze zu riskieren, ist dies ihre Wahl.
Aber minderjährige Kinder unterliegen den Entscheidungen ihrer Eltern, also bitte geben Sie Ihrem Kind KEINE COVID-19-Spritze!
Beachten Sie die Warnung dieser Eltern, die es jetzt bereuen, dass ihre Kinder eine dieser Spritzen bekommen haben, weil ihr Kind jetzt tot ist oder an einer Herzkrankheit leidet .
Teile dieses Video weit und breit. Es ist auf unserem Bitchute-Kanal und Sie können es auch von unserem Telegram-Kanal herunterladen .
2.809 tote Babys in VAERS nach COVID-Schüssen, da neue Dokumente beweisen, dass Pfizer, die FDA und die CDC wussten, dass die Schüsse für schwangere Frauen nicht sicher sind
Alle Artikel von Global Research können in 51 Sprachen gelesen werden, indem das Dropdown-Menü „Website übersetzen“ im oberen Banner unserer Homepage (Desktop-Version) aktiviert wird.
Um den täglichen Newsletter von Global Research (ausgewählte Artikel) zu erhalten, klicken Sie hier .
Besuchen und folgen Sie uns auf Instagram unter @ crg_globalresearch .
***
Der neueste Daten-Dump in das Vaccine Adverse Events Reporting System (VAERS) der US-Regierung erfolgte gestern (12.03.21) und umfasst Daten bis 26.11.2021.
Es werden nun 927.740 Fälle an VAERS gemeldet, die in den letzten 11 Monaten nach COVID-19- Impfungen aufgetreten sind, von den insgesamt 1.782.453 Fällen in der gesamten VAERS-Datenbank, die in den letzten 30+ Jahren erfasst wurden.
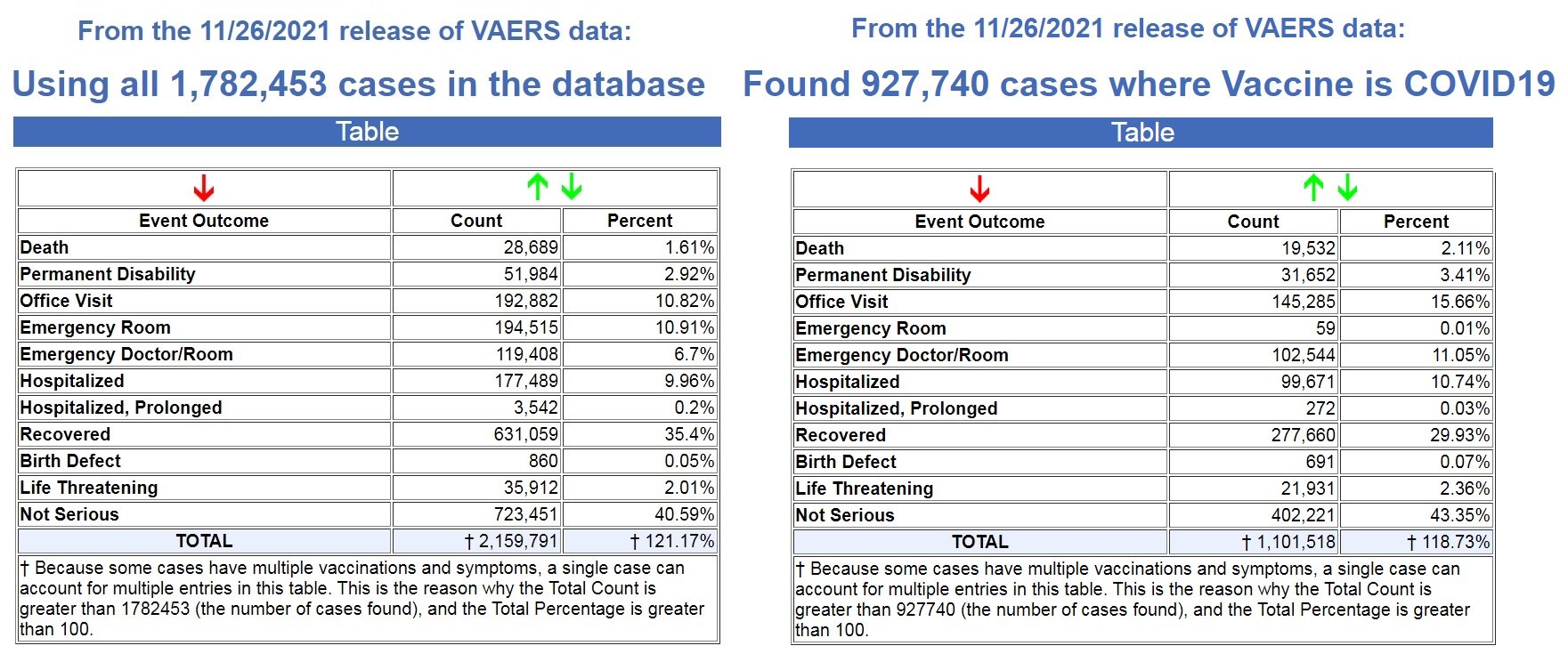
Linke Bildquelle , Rechte Bildquelle .
Das bedeutet, dass in den letzten 11 Monaten nach den COVID-19-Spritzen 52 % aller Fälle von Impfstoff-Nebenwirkungen in VAERS in den letzten 30+ Jahren gemeldet wurden.
Darüber hinaus wurden 68 % aller Todesfälle nach Impfstoffen, die in den letzten 30+ Jahren in VAERS gemeldet wurden, in den letzten 11 Monaten nach den COVID-19-Impfungen gemeldet.
Wir sind auf dem besten Weg , 21.307 Todesfälle im ersten Jahr nach den experimentellen COVID-19-Impfungen zu verzeichnen, während die durchschnittlichen jährlichen Todesfälle nach FDA-zugelassenen Impfstoffen in den letzten 30+ Jahren 305 Todesfälle betragen .
Das ist ein erstaunlicher Anstieg der gemeldeten Todesfälle von 86% nach den COVID-19-Impfungen, ein 70-facher Anstieg gegenüber den durchschnittlich gemeldeten Todesfällen nach Impfungen in den letzten 30+ Jahren!
- FDA-zugelassene Impfstoffe: 305 Todesfälle pro Jahr
- COVID-19 EUA-Schüsse: 21.307 Todesfälle pro Jahr
Und wie Dr. Jessica Rose zuvor berichtet hat , beträgt der Untererfassungsfaktor in VAERS für die COVID-19-Spritzen 41X, als konservative Zahl, was bedeutet, dass basierend auf den VAERS jetzt mindestens 800.812 Menschen nach COVID-19-Spritzen gestorben sind Daten.
Die meisten, wenn nicht alle dieser Todesfälle werden in den pharmaeigenen Unternehmensmedien als „COVID“ -Todesfälle gemeldet, da in den ersten 11 Monaten des Jahres 2021 mehr „COVID-Todesfälle“ registriert wurden als im gesamten Jahr des Jahres 2021 2020, als es bis Dezember keine COVID-Impfstoffe gab. ( Quelle. )
Satire

Rekordzahl fetaler Todesfälle nach COVID-19-Schüssen
Mit dieser neuesten Aktualisierung in VAERS haben wir nun 2.809 fötale Todesfälle nach COVID-19-Spritzen bei schwangeren und gebärenden Frauen in den letzten 11 Monaten gefunden. ( Quelle .)
Im Gegensatz dazu fanden wir unter Verwendung der exakt gleichen Suchparameter in VAERS, aber ohne die COVID-19-Spritzen, 2.168 fötale Todesfälle nach allen von der FDA zugelassenen Impfstoffen in den letzten 30+ Jahren. ( Quelle .)
Das sind durchschnittlich 72 fötale Todesfälle pro Jahr nach allen von der FDA zugelassenen Impfstoffen in den letzten 30+ Jahren, verglichen mit 3064 fötalen Todesfällen in 1 Jahr nach COVID-19-Impfungen.
- FDA-zugelassene Impfstoffe: 72 Todesfälle bei Föten pro Jahr
- COVID-19 EUA-Schüsse: 3064 fötale Todesfälle pro Jahr
Das ist eine 80-prozentige Zunahme der fetalen Todesfälle, die in VAERS nach den COVID-19-Spritzen verzeichnet wurden. Dennoch empfehlen die CDC und die FDA diese EUA-Schüsse weiterhin für schwangere Frauen und stillende Mütter.
Sie empfehlen diese Spritzen nicht nur für schwangere Frauen, wir haben jetzt auch genügend Beweise dafür, dass sie seit Anfang dieses Jahres wissen, dass diese Spritzen für schwangere Frauen gefährlich sind und zum Tod des Fötus führen.
In einer Sitzung der Beratungskommission für Impfstoffe im Kindesalter (ACCV) am 4. März 2021 legte die CDC einen Bericht vor , der einen Abschnitt mit dem Titel: Zusammenfassung der Impfsicherheit für Mütter (ab S. 39) enthielt .
Sie sagten (Hervorhebung von mir – meine Kommentare in Rot):
* Schwangere wurden nicht ausdrücklich in klinische Studien vor der Zulassung von COVID-19-Impfstoffen eingeschlossen
– Sicherheitsüberwachung und -forschung nach der Zulassung sind die wichtigsten Möglichkeiten, um Sicherheitsdaten zur COVID-19-Impfung während der Schwangerschaft zu erhalten
* Größere Zahl von Selbstangaben als erwartet Schwangere haben sich bei v-safe registriert
* Das Reaktogenitätsprofil und die bei Schwangeren bei v-safe beobachteten unerwünschten Ereignisse zeigten keine Sicherheitsprobleme (basierend auf welchen Kriterien???)
* Die meisten Meldungen an VAERS bei Schwangeren (73%) beteiligt nicht schwangerschaftsspezifische unerwünschte Ereignisse (z. B. lokale und systemische Reaktionen)
* Fehlgeburten waren die am häufigsten berichteten schwangerschaftsspezifischen unerwünschten Ereignisse an VAERS; Zahlen liegen innerhalb der bekannten Hintergrundraten, basierend auf mutmaßlichen COVID-19-Impfstoffdosen, die schwangeren Frauen verabreicht wurden (keine stützenden Beweise zur Untermauerung dieser Behauptungen).
Es ist wichtig, bei all diesen Berichten der CDC zu beachten, dass diese auf Selbstberichtsdaten von schwangeren Frauen basieren .
Wir wissen , dass es politisch inkorrekt ist jedes gesundheitliches Problem auf einem COVID-19 schuld „Impfstoff“ , und dass Ärzte und Krankenschwestern sind unter Druck diese nicht zu berichten , so wie viele schwangeren Frauen eine negative Reaktion hatten, wie eine Fehlgeburt, und noch nie dachten, es mit ihrer COVID-19-Aufnahme zu verbinden?
Bereits im März dieses Jahres (2021) gab es daher große Bedenken hinsichtlich der Auswirkungen der Spritzen auf Schwangere, da Schwangere „größer als erwartet“ über Nebenwirkungen berichteten und „die am häufigsten gemeldeten schwangerschaftsspezifischen Nebenwirkungen“ für VAERS“ war „ Fehlgeburt “.
Im August dieses Jahres (2021) präsentierte die CDC dann eine „neue Studie“ mit „neuen Daten“.
Auch hier hängen diese „Daten“ davon ab, dass schwangere Frauen Nebenwirkungen „selbst berichten“. COVID-19-Aufnahmen. Dies zu tun bedeutet, als "Anti-Vaxxer" gebrandmarkt zu werden und dich fürs Leben zu beschämen.
Das Update vom August gab zu, dass 13 % der schwangeren Frauen, die eine COVID-19-Impfung erhalten hatten, eine Fehlgeburt gemeldet hatten. Die CDC wischte dies beiseite, indem sie feststellte, dass "Fehlgeburten typischerweise bei etwa 11-16% der Schwangerschaften auftreten".
Aber natürlich werden ALLE Fehlgeburten irgendwo in der Krankenakte gemeldet, weshalb sie sogar einen solchen Nummernkreis angeben können. Diese Zahl basiert also auf 100% der gemeldeten Daten, während die COVID-19-bezogenen Fehlgeburten nur auf Selbstmeldungen basieren und wir keine Ahnung haben, wie viele Frauen ihre Fehlgeburten nie gemeldet haben, weil sie es nie mit ihrem COVID in Verbindung gebracht haben -19 Schuss.
Eine der Hauptstudien, auf die sich die CDC angeblich stützte, um zu erklären, dass COVID-19-Spritzen für schwangere Frauen sicher sind, war eine Studie, die am 17. Juni 2021 im New England Journal of Medicine veröffentlicht wurde.
Aber am 14. Oktober 2021 gaben sie eine Erklärung ab, dass einige ihrer Daten in der Studie vom 17. Juni falsch waren. ( Quelle .) Es befasste sich speziell mit Schwangerschaften in der 20. Woche oder früher.
„Es stand kein Nenner zur Verfügung, um eine Risikoabschätzung für Spontanaborte zu berechnen, da zum Zeitpunkt dieses Berichts für 905 der 1224 Teilnehmerinnen, die innerhalb von 30 Tagen vor dem ersten Tag der letzten Menstruation geimpft wurden, eine Nachbeobachtung über 20 Wochen noch nicht verfügbar war.“ Periode oder im ersten Trimester. Darüber hinaus müsste jede Risikoabschätzung das in der Schwangerschaftswoche spezifische Risiko eines Spontanaborts berücksichtigen.“ ( Quelle .)
In diesem Video, das wir im Oktober produziert haben, diskutieren Dr. Byram Bridle in Kanada und Dr. Martin Kulldorff von der Harvard Medical School mit Laura Ingraham die Bedeutung dieses Fehlers, der in dieser Studie gemacht wurde, die die CDC-Politik zu Fox News bestimmt hat. (Es ist in der zweiten Hälfte des Videos nach den Beispielen für unerwünschte Ereignisse bei Säuglingen.)
Seitdem haben Forscher in Neuseeland eine neue Studie mit den Originaldaten durchgeführt und kamen zu dem Schluss:
Eine erneute Analyse dieser Zahlen zeigt eine kumulative Inzidenz von Spontanaborten von 82 % (104/127) bis 91 % (104/114), 7–8 Mal höher als die Ergebnisse der ursprünglichen Autoren. ( Quelle .)
Dennoch empfehlen die CDC und die FDA weiterhin die Impfungen für schwangere Frauen, obwohl eine korrekte Analyse der Originaldaten zeigt, dass 82 bis 91 % der schwangeren Frauen Fehlgeburten erleiden, wenn ihr ungeborenes Kind weniger als 20 Wochen alt ist. ( Quelle .)
Wir haben auch Beweise dafür, dass Pfizer über das Risiko ihrer COVID-19-Impfungen für schwangere Frauen Bescheid wusste.
Im Mai dieses Jahres (2021) veröffentlichten wir einen Bericht von Bud Shaver von Abortion Free New Mexico, der auf einem Whistleblower basiert, der in einer COVID-19-Taskforce tätig war und Dokumente gefunden hatte, die Pfizer bei der Europäischen Arzneimittelagentur (EMA) eingereicht hatte ), um die Schüsse in Europa zu genehmigen, wozu auch Tierversuche gehörten, die bei Rattenexemplaren schwerwiegende Geburtsfehler aufwiesen.
In den Pfizer-Factsheets heißt es, dass schwangere oder stillende Frauen ihre Möglichkeiten mit ihrem Gesundheitsdienstleister besprechen sollten. Obwohl die US-amerikanische FDA die vollständigen Studiendetails, die ihnen zur Genehmigung der Notfallzulassung (EUA) zur Verfügung gestellt wurden, nicht veröffentlicht hat, hat die Europäische Arzneimittelbehörde dies getan.
Die vollständigen Studiendokumente sind unter www.ema.europa.eu verfügbar .
Laut der Reproduktionstoxizitätsstudie des Pfizer-Produkts, die an trächtigen Ratten durchgeführt wurde: „ Es gab eine Zunahme (~2x) des Verlusts vor der Implantation (Schwangerschaft)“ und „eine sehr geringe Inzidenz von Gastroschisis, Mund-/Kieferfehlbildungen, richtig“. seitlichen Aortenbogen und Halswirbelanomalien.“
Sie behaupten, dass diese Schwangerschaftsreduktionen innerhalb normaler histologischer Bereiche liegen, sie wurden jedoch konsistent beobachtet und sind wahrscheinlich statistisch signifikant. Bei der Gastroschisis wächst der Darm außerhalb des Körpers.
Rechtsseitiger Aortenbogen bedeutet, dass sich das Herz grundsätzlich in die falsche Richtung gebildet hat (der Aortenbogen sollte auf der linken Seite liegen). ( Quelle .)
Dies würde das unterstützen, was wir in VAERS in Bezug auf „ektopische Schwangerschaften“ nach COVID-19-Impfungen gefunden haben, die in den letzten 30+ Jahren 50-mal häufiger als nach ALLEN Impfstoffen gemeldet wurden. Siehe: VAERS-Daten zeigen 50-mal mehr Eileiterschwangerschaften nach COVID-Impfungen als nach ALLEN Impfstoffen in den letzten 30 Jahren
Letzten Monat, im November 2021, haben wir den Bericht von Rechtsanwalt Aaron Siri veröffentlicht , einem Anwalt für Impfverletzungen, der die FDA im Namen mehrerer Ärzte verklagt, die die Kläger sind und sich entschieden haben, ihre Karriere aufs Spiel zu setzen, um eine Entlarvung zu wagen Impfstofftote und -verletzungen, die durch die experimentellen COVID-19-Schüsse verursacht wurden.
Anwältin Siri schrieb, dass Pfizer beantragt habe, 55 Jahre Zeit zu haben, um ihre Studiendaten zu den COVID-19-Schüssen bereitzustellen.
Die FDA hat einen Bundesrichter gebeten , die Öffentlichkeit bis zum Jahr 2076 warten zu lassen, um alle Daten und Informationen offenzulegen, auf die sie sich bei der Lizenzierung des COVID-19-Impfstoffs von Pfizer stützte. Das ist kein Tippfehler. Sie will 55 Jahre, um diese Informationen der Öffentlichkeit zugänglich zu machen.
Also, lassen Sie uns das klarstellen. Die Bundesregierung schirmt Pfizer von der Haftung ab . Gibt es Milliarden von Dollar . Bringt Amerikaner dazu, sein Produkt zu nehmen . Sie werden jedoch nicht die Daten sehen, die die Sicherheit und Wirksamkeit des Produkts belegen. Für wen arbeitet die Regierung? ( Quelle .)
In einem Folgeartikel, den er am 19. November 2021 veröffentlichte, berichtete er, dass der Richter Pfizer gezwungen habe, mit der Veröffentlichung der Daten zu beginnen, und dass sie die ersten 91+ Seiten veröffentlicht hätten.
Zwei Monate und einen Tag nach der Klage und fast drei Monate nach der Zulassung des Covid-19-Impfstoffs von Pfizer veröffentlichte die FDA die erste Runde von Dokumenten, die sie vor der Zulassung dieses Produkts überprüfte. Die Produktion bestand aus 91 pdf-Seiten, einer xpt-Datei und einer txt-Datei. Sie können sie hier herunterladen .
Während es Sache der Wissenschaftler ist, richtig zu analysieren, möchte ich eine Beobachtung teilen. Eines der erstellten Dokumente ist eine kumulative Analyse der Berichte über unerwünschte Ereignisse nach der Zulassung von [dem Impfstoff] , die bis zum 28. Februar 2021 erhalten wurden , was nur 2 ½ Monate nach Erhalt der Notfallgenehmigung (EUA) des Impfstoffs ist. Dieses Dokument spiegelt unerwünschte Ereignisse nach Impfungen wider, die den „Workflow-Zyklus“ von Pfizer innerhalb und außerhalb der USA bis zum 28. Februar 2021 abgeschlossen haben.
Pfizer erklärt auf Seite 6, dass „Aufgrund der großen Anzahl spontaner Berichte über unerwünschte Ereignisse, die für das Produkt eingegangen sind, [Pfizer] der Bearbeitung schwerwiegender Fälle Priorität eingeräumt hat …“ und dass Pfizer „auch [sic] mehrere Maßnahmen ergriffen hat, um helfen, die starke Zunahme von Meldungen über unerwünschte Ereignisse zu mildern“, einschließlich „Erhöhung der Zahl der Kollegen bei der Dateneingabe und Fallbearbeitung“ und „hat ungefähr [ANONYMISIERT] zusätzliche Vollzeitmitarbeiter (FTE) an Bord genommen.“ Fragen Sie, warum es urheberrechtlich geschützt ist, mitzuteilen, wie viele Personen Pfizer einstellen musste, um alle unerwünschten Ereignisse zu verfolgen, die kurz nach der Einführung des Produkts gemeldet wurden.
Was den Umfang der Meldungen betrifft, so erhielt Pfizer in den 2 ½ Monaten nach der EUA insgesamt 42.086 Meldungen mit 158.893 „Ereignissen“. Die meisten dieser Berichte stammten aus den USA und betrafen überproportional Frauen (29.914 vs. 9.182 von Männern) und Personen zwischen 31 und 50 Jahren (13.886 vs. 21.325 für alle anderen Altersgruppen zusammengenommen, weitere 6.876, deren Alter unbekannt war). Außerdem wurden 25.957 der Ereignisse als „Erkrankungen des Nervensystems“ klassifiziert. ( Quelle .)
Ende Februar dieses Jahres (2021), als Pfizer bei der FDA die vollständige Zulassung ihrer EUA-COVID-19-Spritze beantragte, lagen ihnen bereits Daten aus 42.086 Berichten vor, die 158.893 „Ereignisse“ enthielten, von denen Frauen im Alter von überproportional betroffen waren 31 und 50.
In dem hier zu findenden „Postmarketing“-Bericht von Pfizer gibt es eine Tabelle, Tabelle 6 mit der Bezeichnung „Beschreibung fehlender Informationen“ für „Anwendung in Schwangerschaft und Stillzeit“, die 274 Fälle abdeckt und besagt:
Schwangerschaftsfälle: 274 Fälle, darunter:
• 270 Fälle von Müttern und 4 Fälle von Fötus/Baby, die 270 einzigartige Schwangerschaften repräsentieren (die 4 Fälle von Fötus/Baby wurden mit 3 Fällen von Müttern in Verbindung gebracht; 1 Fall von Mutter betraf Zwillinge).
• Die Schwangerschaftsergebnisse der 270 Schwangerschaften wurden als Spontanabort (23), Outcome ausstehend (5), Frühgeburt mit Tod des Neugeborenen, Spontanabort mit intrauterinem Tod (je 2), Spontanabort mit Tod des Neugeborenen und normales Ergebnis (je 1 .) berichtet ). Für 238 Schwangerschaften wurde kein Ergebnis angegeben (beachten Sie, dass für jeden Zwilling 2 verschiedene Ergebnisse gemeldet und beide gezählt wurden).
• 146 nicht schwerwiegende Fälle von Müttern berichteten über eine Impfstoffexposition in utero, ohne dass ein klinisches unerwünschtes Ereignis aufgetreten war. Die Expositions-PTs kodiert zu den PTs Mütterliche Exposition während der Schwangerschaft (111), Exposition während der Schwangerschaft (29) und mütterlicher Expositionszeitpunkt nicht spezifiziert (6). In 21 dieser Fälle wurde über ein Trimester der Exposition berichtet: 1. Trimester (15 Fälle), 2. Trimester (7) und 3. Trimester (2).
• 124 Fälle von Müttern, 49 nicht schwerwiegende und 75 schwere, berichtete klinische Ereignisse, die bei den geimpften Müttern auftraten. Schwangerschaftsbezogene Ereignisse, über die in diesen Fällen berichtet wurde, kodiert mit den PTs Abort spontan (25), Uteruskontraktion während der Schwangerschaft, vorzeitiger Blasensprung, Abort, Fehlgeburt und fetaler Tod (jeweils 1).
• 4 schwere Fötus-/Baby-Fälle berichteten über die PTs Exposition während der Schwangerschaft, fetale Wachstumseinschränkung, mütterliche Exposition während der Schwangerschaft, Frühgeborene (jeweils 2) und Tod des Neugeborenen (1). Für 2 Fälle (Zwillinge) wurde während des 1. Trimesters der Exposition berichtet.
Dies waren die Daten, die die FDA verwendet hat, um den Pfizer COVID-19-Schuss zu genehmigen.
Sie stellten der FDA auch Daten für stillende Babys zur Verfügung, die eindeutig darauf hindeuteten, dass die Schüsse diese Babys betrafen.
Fälle
beim Stillen von Säuglingen: 133, davon: • 116 Fälle, in denen eine Impfstoffexposition während des Stillens (PT-Exposition über die Muttermilch) ohne das Auftreten klinischer Nebenwirkungen berichtet wurde;
• 17 Fälle, 3 schwerwiegende und 14 nicht schwerwiegende, berichteten über die folgenden klinischen Ereignisse, die bei Säuglingen/Kind, die über das Stillen dem Impfstoff ausgesetzt wurden, auftraten: Fieber (5), Hautausschlag (4), Reizbarkeit des Säuglings (3), Erbrechen bei Säuglingen, Durchfall , Schlaflosigkeit und Krankheit (jeweils 2), schlechte Ernährung des Säuglings, Lethargie, Bauchbeschwerden, Erbrechen, Impfstoffallergie, gesteigerter Appetit, Angst,
Weinen, schlechte Schlafqualität, Aufstoßen, Erregung, Schmerzen und Urtikaria (je 1).
Fälle von stillenden Müttern (6):
• 1 schwerwiegender Fall berichtet 3 klinische Ereignisse, die bei einer Mutter während des Stillens auftraten (PT Mütterliche Exposition während des Stillens); diese Ereignisse wurden mit den PTs Schüttelfrost,
Unwohlsein und Fieber kodiert • 1 nicht schwerwiegender Fall mit sehr begrenzten Informationen und ohne assoziierte UE berichtet. ( Quelle .)
Und das war Ende Februar. Wir können deutlich sehen, welche Ergebnisse seitdem bei ungeborenen Kindern erzielt wurden, nur basierend auf den begrenzten Daten, die an VAERS gemeldet wurden, wo nach den COVID-19-Spritzen ein Anstieg der fötalen Todesfälle in VAERS um 80 % zu verzeichnen war.
Jetzt bin ich nur noch ein Reporter, der hinter einem Computer sitzt und auf diese öffentlich verfügbaren Daten zugreift, damit ich es Ihnen melden kann.
Sie können sicher sein, dass auch die Wissenschaftler und Forscher von Pfizer, der FDA und der CDC Zugriff auf all diese Daten haben.
Allein dieser Artikel mit all den Links zu den öffentlich zugänglichen Daten enthält mehr als genug Informationen, um sofort Haftbefehle gegen Rochelle Walensky, die Direktorin der CDC, Janet Woodcock, die Direktorin der FDA, und Albert Bourla, den CEO von Pfizer, auszustellen. wegen Massenmord und Verbrechen gegen die Menschlichkeit.
Aber gibt es irgendwo in den Vereinigten Staaten einen Anwalt, der diese Haftbefehle ausstellen würde?
Wir können Bidens Generalstaatsanwalt für die USA, Merrick Garland, ziemlich ausschließen.
Gibt es in den 50 Vereinigten Staaten einen Generalstaatsanwalt, der den Mut und den Segen ihres Gouverneurs hätte, Haftbefehle wie diesen zu erlassen?
Unwahrscheinlich, da kein einziger Gouverneur eines Staates, ob rot oder blau, Maßnahmen ergriffen hat, um Leben zu schützen und die Kriminellen hinter diesen Biowaffenschüssen zu verhaften.
Da es sich jedoch um Bundesbehörden, die FDA und die CDC, handelt, die jeden einzelnen Bürger der Vereinigten Staaten betreffen, könnte ein Bezirksstaatsanwalt Haftbefehle ausstellen und versuchen, ihnen zu dienen. Sie würden höchstwahrscheinlich so etwas wie eine Milizgruppe brauchen, die vielleicht aus Sheriff-Abgeordneten und Mitgliedern ihrer Nationalgarde besteht, um so etwas versuchen zu können.
Aber wenn überhaupt nichts unternommen wird, werden diese Todesfälle weiter ansteigen, da sie jetzt Kindern zwischen 5 und 11 Jahren injizieren und sich darauf vorbereiten, Babys im Alter zwischen 6 Monaten und 4 Jahren bald zu injizieren. kurz nach dem ersten des Jahres.
Sind dies die USA, in denen Sie leben und ein Teil davon sein möchten? Wie lange werden wir zusehen, wie unschuldige Menschen getötet werden, um die eugenischen Pläne der Globalisten zur Reduzierung unserer Bevölkerung zu erfüllen?
Wenn in der giftigen RNA Impfung Graphenoxyd ist, wirkt es wie folgt
Erste umfassende Untersuchung von Graphenoxyd aus dem Jahre 2016 nach Entdeckung 2004
Erst-Veröffentlicht: 31. Oktober 2016 Luckenwalde den 06.12. 2021
Toxizität von Nanopartikeln der Graphenfamilie: ein allgemeiner Überblick über die Ursprünge und Mechanismen Forscher: Lingling Ou, Bin Song, Huimin Liang, Jia Liu, Xiaoli Feng, Bin Deng, Ting Sun & Longquan Shao
Abstrakt
Aufgrund ihrer einzigartigen physikalisch-chemischen Eigenschaften werden Nanopartikelmaterialien der Graphen-Familie (GFNs) in vielen Bereichen, insbesondere in biomedizinischen Anwendungen, verwendet.
Derzeit haben viele Studien die Biokompatibilität und Toxizität von GFNs in vivo und intro untersucht.
Im Allgemeinen können GFNs in Tieren oder Zellmodellen unterschiedliche Toxizitätsgrade ausüben, indem sie verschiedenen Verabreichungswegen folgen und physiologische Barrieren durchdringen, anschließend in Geweben verteilt oder in Zellen lokalisiert werden und schließlich aus dem Körper ausgeschieden werden.
Dieser Review sammelt Studien zu den toxischen Wirkungen von GFNs in verschiedenen Organen und Zellmodellen.
Wir weisen auch darauf hin, dass verschiedene Faktoren die Toxizität von GFNs bestimmen, einschließlich der lateralen Größe, Oberflächenstruktur, Funktionalisierung, Ladung, Verunreinigungen, Aggregationen und Koronaeffekt ect.
Darüber hinaus wurden mehrere typische Mechanismen aufgedeckt, die der GFN-Toxizität zugrunde liegen, beispielsweise physische Zerstörung, oxidativer Stress, DNA-Schäden, Entzündungsreaktionen, Apoptose, Autophagie und Nekrose.
An diesen Mechanismen sind (toll-like Rezeptoren-) TLR-, Transforming Growth Factor β- (TGF-β-) und Tumornekrosefaktor-alpha (TNF-α) abhängige Pfade im Signalweg-Netzwerk sowie oxidativer Stress beteiligt spielt auf diesen Wegen eine entscheidende Rolle. In diesem Review fassen wir die verfügbaren Informationen zu regulierenden Faktoren und den Mechanismen der GFN-Toxizität zusammen und schlagen einige Herausforderungen und Vorschläge für weitere Untersuchungen von GFNs vor, mit dem Ziel, die toxikologischen Mechanismen zu vervollständigen und Vorschläge zur Verbesserung der biologischen Sicherheit von GFNs zu machen und erleichtern deren breite Anwendung.
Hintergrund
Graphen, das aus kristallinem Graphit isoliert wird, ist eine flache Monoschicht, die aus einatomigen dicken, zweidimensionalen Blättern eines hexagonal angeordneten Wabengitters besteht [1].
Aufgrund seiner einzigartigen strukturellen, spezifischen Oberfläche und mechanischen Eigenschaften haben die Funktionen und Anwendungen von Graphen seit der Entdeckung des Materials im Jahr 2004 große Aufmerksamkeit erlangt [2, 3]. Graphen und seine Derivate umfassen Monolayer-Graphen, Wenigschicht-Graphen (FLG), Graphenoxid (GO), reduziertes Graphenoxid (rGO), Graphen-Nanoblätter (GNS) und Graphen-Nanobänder usw. [4–7].
GO ist eines der wichtigsten chemischen Graphen-Derivate der Graphen-Familie von Nanomaterialien (GFNs), das aufgrund seiner potenziellen biomedizinischen Anwendungen immer mehr Aufmerksamkeit auf sich zieht. Graphenbasierte Materialien haben normalerweise Größen von einigen bis zu Hunderten von Nanometern und sind 1-10 nm dick [8, 9], was auch die Definition von „Nanopartikeln“ oder „Nanomaterialien“ ist. Aufgrund ihrer außergewöhnlichen physikalischen und chemischen Eigenschaften werden Graphenmaterialien in verschiedenen Bereichen weit verbreitet verwendet, einschließlich der Energiespeicherung; nanoelektronische Geräte; Batterien [10-12]; und biomedizinische Anwendungen, wie antibakterielle Mittel [13, 14], Biosensoren [15–18], Zellbildgebung [19, 20], Wirkstoffabgabe [8, 21, 22] und Gewebezüchtung [23–25].
Mit zunehmender Anwendung und Produktion von GFNs steigt auch das Risiko einer unbeabsichtigten beruflichen oder umweltbedingten Exposition gegenüber GFNs [26]. Und vor kurzem gab es einige Untersuchungen zur GFN-Exposition in beruflichen Umgebungen, und veröffentlichte Daten zeigten, dass die berufliche Exposition von GFNs eine potenzielle Toxizität für Arbeiter und Forscher hatte [27–29]. GFNs können dem Körper durch intratracheale Instillation [30], orale Verabreichung [31], intravenöse Injektion [32], intraperitoneale Injektion [33] und subkutane Injektion [34] zugeführt werden.
GFNs können akute und chronische Gewebeverletzungen hervorrufen, indem sie die Blut-Luft-Schranke, die Blut-Hoden-Schranke, die Blut-Hirn-Schranke und die Blut-Plazenta-Schranke usw. durchdringen und sich in Lunge, Leber und Milz usw. anreichern.
Einige Graphen-Nanomaterialien-Aerosole können eingeatmet werden und eine erhebliche Ablagerung in den Atemwegen verursachen, und sie können leicht durch die tracheobronchialen Atemwege eindringen und dann in die unteren Atemwege der Lunge gelangen, was zur nachfolgenden Bildung von Granulomen, Lungenfibrose und nachteiligen gesundheitlichen Auswirkungen bei Exposition führt Personen [2, 29]. Mehrere Übersichten haben die einzigartigen Eigenschaften skizziert [35, 36] und die neuesten potentiellen biologischen Anwendungen von GFNs für die Wirkstoffabgabe, Genabgabe, Biosensoren, Gewebezüchtung und Neurochirurgie zusammengefasst [37–39]; bewertete die Biokompatibilität von GFNs in Zellen (Bakterien, Säugetiere und Pflanzen) [7, 40, 41] und Tieren (Mäuse und Zebrafische) [42]; sammelten Informationen über den Einfluss von GFNs in der Boden- und Wasserumgebung [43].
Obwohl in diesen Übersichten die entsprechenden Sicherheitsprofile und die Nanotoxikologie von GFNs erörtert wurden, waren die spezifischen Schlussfolgerungen und detaillierten Toxizitätsmechanismen unzureichend, und die Toxizitätsmechanismen wurden nicht vollständig zusammengefasst.
Die in neueren Studien nachgewiesenen toxikologischen Mechanismen von GFNs umfassen hauptsächlich Entzündungsreaktionen, DNA-Schäden, Apoptose, Autophagie und Nekrose usw., und diese Mechanismen können gesammelt werden, um das komplexe Netzwerk der Signalwege, das die Toxizität von GFNs reguliert, weiter zu erforschen. Es muss darauf hingewiesen werden, dass es mehrere Faktoren gibt, die die Toxizität von GFNs stark beeinflussen, wie z der Toxizität von GFNs in vitro und in vivo durch verschiedene experimentelle Methoden, mit dem Ziel, Vorschläge für weitere Studien von GFNs zu geben und die toxikologischen Mechanismen zu vervollständigen, um die biologische Sicherheit von GFNs zu verbessern und ihre breite Anwendung zu erleichtern.
Toxizität von GFNs (in vivo und in vitro)
GFNs durchdringen die physiologischen Barrieren oder Zellstrukturen durch verschiedene Expositionswege oder Verabreichungswege und dringen in den Körper oder die Zellen ein, was schließlich zu einer Toxizität in vivo und in vitro führt. Die unterschiedlichen Verabreichungswege und Eintrittswege, die unterschiedliche Gewebeverteilung und Ausscheidung, sogar die unterschiedlichen Aufnahmemuster und -orte der Zellen können den Grad der Toxizität von GFNs bestimmen [44–46]. Sie zu verdeutlichen, kann daher hilfreich sein, um die Gesetze des Auftretens und der Entwicklung der GFN-Toxizität besser zu verstehen.
Verwaltungsweg
Die üblichen Verabreichungswege in Tiermodellen umfassen Atemwegsexposition (intranasale Insufflation, intratracheale Instillation und Inhalation), orale Verabreichung, intravenöse Injektion, intraperitoneale Injektion und subkutane Injektion. Der Hauptexpositionsweg für GFNs in der Arbeitsumgebung ist die Atemwegsexposition, daher werden Inhalation und intratracheale Instillation hauptsächlich bei Mäusen verwendet, um die Exposition des Menschen gegenüber GFNs zu simulieren. Obwohl die Inhalationsmethode die realistischste Simulation der Exposition im wirklichen Leben bietet, ist die Instillation eine effektivere und zeitsparendere Methode, und es wurde festgestellt, dass GFNs eine längere Entzündungsperiode unter Instillation (intratracheale Instillation, intrapleurale Installation und pharyngeale Aspiration) als Inhalation verursachen [24, 30, 47, 48]. Es wurde untersucht, dass sich GFNs in der Lunge ablagern und zu einem hohen Niveau akkumulieren, das nach intratrachealer Instillation für mehr als 3 Monate in der Lunge mit langsamer Klärung verbleibt [49]. Die intravenöse Injektion wird auch häufig verwendet, um die Toxizität von Graphen-Nanomaterialien zu bewerten, und Graphen zirkuliert innerhalb von 30 Minuten durch den Körper von Mäusen und reichert sich in einer Arbeitskonzentration in Leber und Blase an [32, 50–52]. GO-Derivate hatten jedoch eine eher endliche intestinale Adsorption und wurden bei erwachsenen Mäusen durch orale Verabreichung schnell ausgeschieden [31, 53]. Nanogroßes GO (350 nm) führte dazu, dass nach subkutaner Injektion im Nackenbereich weniger mononukleäre Zellen in das subkutane Fettgewebe infiltrierten als mikrometergroßes GO (2 μm) [34]. GO agglomerierte nach intraperitonealer Injektion in der Nähe der Injektionsstelle, und zahlreiche kleinere Aggregate setzten sich in der Nähe der Leber- und Milzserosa ab [31, 33]. Experimente zu Hautkontakt mit oder Hautpermeation von GFNs wurden in den hier besprochenen Veröffentlichungen nicht gefunden, und es liegen keine ausreichenden Beweise vor, um den Schluss zu ziehen, dass Graphen intakte Haut oder Hautläsionen durchdringen kann. Der Weg der Nasentropfen, der häufig verwendet wurde, um die Neurotoxizität oder das Potenzial anderer Nanomaterialien für Hirnschädigungen zu testen, wurde in den hier besprochenen Artikeln nicht erwähnt.
Zugangspfade für GFNs
GFNs gelangen nach dem Eintritt in den Körper durch den Blutkreislauf oder durch biologische Barrieren an verschiedene Stellen, was zu einer unterschiedlichen Retention in verschiedenen Organen führt. Aufgrund ihrer Nanogröße können GFNs tiefere Organe erreichen, indem sie die normalen physiologischen Barrieren wie die Blut-Luft-Schranke, die Blut-Hoden-Schranke, die Blut-Hirn-Schranke und die Blut-Plazenta-Schranke passieren.
Blut-Luft-Schranke
Die Lunge ist ein potenzieller Eingang für Graphen-Nanopartikel in den menschlichen Körper durch die Atemwege. Die inhalierten GO-Nanoblätter können die Ultrastruktur und die biophysikalischen Eigenschaften des pulmonalen Tensidfilms (PS) zerstören, der die erste Verteidigungslinie des Wirts darstellt, und ihre potenzielle Toxizität hervorbringen [54]. Die agglomerierten oder dispergierten Partikel lagern sich auf der inneren Alveolaroberfläche innerhalb der Alveolen ab und werden dann von Alveolarmakrophagen (AMs) verschlungen [55]. Die Clearance in der Lunge wird durch die mukoziliäre Rolltreppe, AMs oder die Epithelschicht erleichtert [56–58]. Einige kleine, inhalierte Nanopartikel infiltrieren jedoch die intakte Lungenepithelbarriere und können dann vorübergehend in das Alveolarepithel oder das Interstitium gelangen [59, 60]. Intratracheal instilliertes Graphen kann sich durch Passieren der Luft-Blut-Schranke in Leber und Milz umverteilen [61]. Die Untersuchung der Blut-Luft-Schranke kann eine intensive Aufmerksamkeit auf sich ziehen, da Forscher und Arbeiter beruflich GFNs in der Regel durch Inhalation ausgesetzt sind. Die Rolle der Blut-Luft-Schranke bei der Toxizität von GFNs zu verdeutlichen, könnte ein heißes Forschungsthema werden.
Blut-Hirn-Schranke
Die komplizierte Anordnung der Blut-Hirn-Schranke, bestehend aus einer Vielzahl von Membranrezeptoren und hochselektiven Trägern, hat im Vergleich zum peripheren Gefäßendothel nur einen geringen Einfluss auf die Durchblutung und die Mikroumgebung des Gehirns [62]. Die Erforschung des Mechanismus der Blut-Hirn-Schranke hatte einige Fortschritte bei Krankheiten und Nanotoxizität gemacht. Matrix-assisted Laser Desorption/Ionization (MALDI)-Massenspektrometrie-Bildgebung (MSI) zeigte, dass rGO mit einem durchschnittlichen Durchmesser von 342 ± 23,5 nm durch den parazellulären Weg in den interendothelialen Spalt zeitabhängig durchdringt, indem die parazelluläre Enge der Blut-Hirn-Schranke [63]. Darüber hinaus können Graphen-Quantenpunkte (GQDs) mit einer geringen Größe von weniger als 100 nm die Blut-Hirn-Schranke passieren [64]. Studien darüber, wie Graphenmaterialien die Blut-Hirn-Schranke passieren und Neurotoxizität verursachen, sind sehr selten, und es sind weitere Daten erforderlich, um eine Schlussfolgerung zu ziehen.
Blut-Hoden-Schranke
Die Blut-Hoden- und Blut-Nebenhoden-Schranken sind dafür bekannt, dass sie zu den engsten Blut-Gewebe-Schranken im Körper von Säugetieren gehören [65]. GO-Partikel mit Durchmessern von 54,9 ± 23,1 nm hatten nach intraabdominaler Injektion Schwierigkeiten, die Blut-Hoden- und Blut-Nebenhoden-Schranke zu durchdringen, und die Spermienqualität der Mäuse war selbst bei einer Dosierung von 300 mg/kg offensichtlich nicht beeinträchtigt [66].
Blut-Plazenta-Schranke
Die Plazentaschranke ist für die Aufrechterhaltung der Schwangerschaft unverzichtbar, da sie den Austausch von Nährstoffen und Stoffwechselschlacken vermittelt, lebenswichtige Stoffwechselfunktionen ausübt und Hormone ausschüttet [67]. Eine kürzlich durchgeführte Überprüfung ergab, dass die Plazenta keine enge Barriere gegen die Übertragung von Nanopartikeln auf Föten darstellt, insbesondere gegen die Verteilung von kohlenstoffhaltigen Nanopartikeln auf und in den Föten [42]. Es wurde vermutet, dass rGO und Goldpartikel (Durchmesser 13 nm) in der späten Schwangerschaft nach intravenöser Injektion in der Plazenta und im Fetus kaum vorhanden sind oder fehlen [44, 68]. Andere Berichte zeigten jedoch, dass ein transplazentarer Transfer in späten Gestationsstadien stattfindet [69, 70]. Der Entwicklungstoxizität von Nanomaterialien wurde viel Aufmerksamkeit gewidmet, und Berichte zeigten, dass viele Nanopartikel die Plazentaschranke passieren und die Entwicklung von Embryonen stark beeinflussten [71–75]. Studien zur Exposition gegenüber Graphenmaterialien durch die Plazentaschranke sind jedoch mangelhaft, und wie diese Partikel auf Embryonen übertragen werden, sollte in Zukunft im Detail evaluiert werden.
Diese vier Barrieren waren die in der Literatur am häufigsten genannten Barrieren, und andere Barrieren wurden in neueren Studien nicht bewertet, wie beispielsweise Hautbarrieren, die in keiner der Hunderte von durchsuchten GFN-Toxizitätsstudien erwähnt wurden. Darüber hinaus ist der Mechanismus, mit dem GFNs diese Barrieren passieren, nicht gut verstanden, und es sind dringend systematischere Untersuchungen erforderlich.
Verteilung und Ausscheidung von GFNs im Gewebe
Die Absorption, Verteilung und Ausscheidung von Graphen-Nanopartikeln kann durch verschiedene Faktoren beeinflusst werden, einschließlich der Verabreichungswege, physikalisch-chemischen Eigenschaften, Partikelagglomeration und Oberflächenbeschichtung von GFNs.
Die unterschiedlichen Verabreichungswege beeinflussen die Verteilung von GFNs, beispielsweise intratracheal instilliertes FLG, das die Luft-Blut-Schranke passiert, hauptsächlich akkumuliert und in der Lunge zurückbleibt, wobei 47 % nach 4 Wochen verbleiben [61]. Intravenös verabreichtes GO gelangte über den Blutkreislauf in den Körper und wurde in Lunge, Leber, Milz und Knochenmark stark zurückgehalten, und in den Lungen von Mäusen wurden nach intravenöser Injektion von 10 mg kg/Körper entzündliche Zellinfiltration, Granulombildung und Lungenödem beobachtet Gewicht GO [49]. In ähnlicher Weise wurde eine hohe Akkumulation von PEGylierten GO-Derivaten im retikuloendothelialen (RES) System einschließlich Leber und Milz nach intraperitonealer Injektion beobachtet. Im Gegensatz dazu zeigten GO-PEG und FLG bei oraler Gabe keine nachweisbare Aufnahme in den Magen-Darm-Trakt oder Gewebeaufnahme [31].
Die unterschiedlichen Eigenschaften von GFNs, wie ihre Größe, Dosis und funktionelle Gruppen, führen immer zu inkonsistenten Ergebnissen in den Verteilungsprofilen von Graphen. Zhang et al. fanden heraus, dass GO hauptsächlich in der Mauslunge eingeschlossen war [49]; Li et al. beobachteten, dass GO in Mäuseleber akkumuliert [76]. Bemerkenswerterweise wurden kleine GO-Faltblätter mit Durchmessern von 10–30 nm hauptsächlich in Leber und Milz verteilt, während sich größere GO-Faltblätter (10–800 nm) hauptsächlich in der Lunge anreicherten [49, 52, 77]. Wenn die Größe von GO größer als die Größe der Gefäße ist, bleibt GO normalerweise in den Arterien und Kapillaren in der Nähe der Injektionsstelle stecken. Es wurde gezeigt, dass die Akkumulation von GO in der Lunge mit einer Erhöhung der injizierten Dosis und Größe zunimmt, die in der Leber jedoch signifikant abnimmt [78]. Die Beschichtung von biokompatiblen Polymeren auf GO beeinflusst auch die Bioverteilung, beispielsweise reichert sich die intravenöse Injektion von GO-PEG und GO-Dextran (GO-DEX) im retikuloendothelialen System (RES), einschließlich Leber und Milz, ohne kurzfristige Toxizität an [ 31, 79]. Darüber hinaus beeinflusst auch die Ladung von Plasmaproteinen und die Adsorption von GO durch Plasmaproteine die Bioverteilung [34].
Die Ausscheidung und Clearance von GFNs variiert in verschiedenen Organen. In der Lunge deuteten Beobachtungen darauf hin, dass NGO von AMs angezogen und beseitigt werden, die durch mukoziliäre Clearance oder auf andere Weise aus dem Sputum eliminiert werden könnten [57] und 46,2 % des intratracheal instillierten FLG wurden 28 d nach der Exposition über den Stuhl ausgeschieden [61]. In der Leber können Nanopartikel über den hepatobiliären Weg nach dem Gallengang in den Zwölffingerdarm eliminiert werden [80]. Darüber hinaus kann PEGyliertes GNS, das sich hauptsächlich in Leber und Milz anreichert, allmählich eliminiert werden, wahrscheinlich sowohl durch die renale als auch durch die fäkale Ausscheidung. Wie kürzlich besprochen, werden GO-Folien größer als 200 nm durch physikalische Milzfiltration abgefangen, aber kleine Größen (ca. 8 nm) können die Nierentubuli in den Urin durchdringen und ohne offensichtliche Toxizität schnell entfernt werden [81]. Die Ausscheidungswege von GFNs sind noch nicht eindeutig geklärt, aber die renalen und fäkalen Wege scheinen die Haupteliminationswege für Graphen zu sein.
In letzter Zeit ist die Verteilungs- und Ausscheidungs-/Toxizitätsstrategie zu einem wichtigen Bestandteil nanotoxikologischer Studien geworden. Bis heute wurden in mehreren Veröffentlichungen mehrere umstrittene Ergebnisse zur Verteilung und Ausscheidung von Graphen in vivo berichtet, und eine systematische Bewertung der Toxikokinetik von GFNs ist noch erforderlich. Der Metabolismus und die Ausscheidung von Nanomaterialien sind lang anhaltende Prozesse, jedoch waren die jüngsten Studien zu GFNs auf kurzfristige toxikologische Bewertungen beschränkt, und die langfristige Akkumulation und Toxizität von GFNs auf verschiedenen Geweben bleibt unbekannt. Daher müssen Langzeitstudien zur Ablagerung und Ausscheidung von GFNs mit verschiedenen Zellen und Tieren durchgeführt werden, um die Biosicherheit der Materialien vor der Verwendung in humanbiomedizinischen Anwendungen zu gewährleisten.
Aufnahme und Lokalisierung von GFNs in Zellen
Es wurde auch beobachtet, dass die Aufnahme und Lokalisation von GFNs in verschiedenen Zelllinien unterschiedliche Wirkungen ausübt. Graphen wird über verschiedene Wege in Zellen aufgenommen [82, 83]. Grundsätzlich sind die physikalisch-chemischen Parameter wie Größe, Form, Beschichtung, Ladung, hydrodynamischer Durchmesser, isoelektrischer Punkt und pH-Gradient wichtig, damit GO die Zellmembran passieren kann [84]. Wie bereits erwähnt, können Nanopartikel mit Durchmessern <100 nm in Zellen eindringen und solche mit Durchmessern <40 nm in den Zellkern [85]. Beispielsweise durchdringen GQDs möglicherweise Zellmembranen direkt und nicht über energieabhängige Wege [86, 87]. Größere proteinbeschichtete Graphenoxid-Nanopartikel (PCGO) (~1 μm) dringen hauptsächlich durch Phagozytose in Zellen ein, und kleinere PCGO-Nanopartikel (~500 nm) gelangen hauptsächlich durch Clathrin-vermittelte Endozytose in Zellen [88]. GO-Faltblätter könnten an der Zellmembran anhaften und umhüllen, sich in die Lipiddoppelschicht einfügen oder als Folge von Interaktionen mit Zellen in die Zelle internalisiert werden [89]. In ähnlicher Weise wurde gezeigt, dass PEGyliertes reduziertes Graphenoxid (PrGO) und rGO aufgrund der Wechselwirkung von hydrophoben, unmodifizierten graphitischen Domänen mit der Zellmembran prominent an der Lipiddoppelschicht-Zellmembran haften [90, 91]. Folglich wurde vermutet, dass eine längere Exposition gegenüber Graphen oder eine hohe Konzentration von Graphen eine physikalische oder biologische Schädigung der Zellmembran sowie eine Destabilisierung der Aktinfilamente und des Zytoskeletts induziert [92].
Aktuelle Daten zeigen, dass GO-Faltblätter mit der Plasmamembran interagieren und von Makrophagen phagozytiert werden. An der Phagozytose des GNS sind drei Hauptrezeptoren auf Makrophagen beteiligt: der Fcg-Rezeptor (FcgR), der Mannose-Rezeptor (MR) und der Komplement-Rezeptor (CR). Darüber hinaus ist FcgR ein Schlüsselrezeptor im vermittelten Phagozytoseweg [90, 93, 94]. Die Proteinkorona von GO fördert die Erkennung durch Makrophagenrezeptoren, insbesondere das in der Proteinkorona enthaltene IgG. Es wurde beobachtet, dass Makrophagen bei Kontakt mit GO erstaunliche morphologische Veränderungen durchmachen [34]. Nach der Internalisierung akkumulierte Graphen im Zellzytoplasma, im perinuklearen Raum und im Zellkern, was bei murinen Makrophagen durch Erhöhung des intrazellulären ROS durch Erschöpfung des mitochondrialen Membranpotentials und durch Auslösen der Apoptose durch Aktivierung des mitochondrialen Signalwegs eine Zytotoxizität induziert [83]. Die möglichen Wechselwirkungen und Akkumulationsstellen von GFNs sind in Abb. 1 zusammengefasst.
Abb. 1
Aus: Toxizität von Nanopartikeln der Graphenfamilie: ein allgemeiner Überblick über die Ursprünge und Mechanismen
Abb. 1
Graphenmaterialien und ihre biologischen Wechselwirkungen. (A) Ein Parameterraum für die am häufigsten verwendeten Graphenmaterialien kann durch die Abmessungen und die Oberflächenfunktionalisierung des Materials beschrieben werden, letztere definiert als der Prozentsatz der Kohlenstoffatome bei der sp3-Hybridisierung. Grüne Quadrate repräsentieren epitaktisch gewachsenes Graphen; gelbes, mechanisch abgeblättertes Graphen; rotes, chemisch abgeblättertes Graphen; blau, Graphenoxid. Beachten Sie, dass in Experimenten auch eine Reihe anderer mit Graphen verwandter Materialien (wie Graphen-Quantenpunkte und Graphen-Nanobänder) verwendet werden. (B) Mögliche Wechselwirkungen zwischen Graphen-verwandten Materialien mit Zellen (die Graphenflocken sind nicht maßstabsgetreu). (a) Adhäsion an der äußeren Oberfläche der Zellmembran. (b) Einbau zwischen die Monoschichten der Lipiddoppelschicht der Plasmamembran. (c) Translokation der Membran. (d) Zytoplasmatische Internalisierung. (e) Clathrin-vermittelte Endozytose. (f) Endosomale oder phagosomale Internalisierung. (g) Lysosomale oder andere perinukleäre Kompartiment-Lokalisierung. (h) Exosomale Lokalisation. Die biologischen Folgen solcher Wechselwirkungen können je nach Kontext der jeweiligen biomedizinischen Anwendung als nachteilig oder vorteilhaft angesehen werden. Verschiedene mit Graphen verwandte Materialien werden unterschiedliche bevorzugte Mechanismen der Interaktion mit Zellen und Geweben aufweisen, die weitgehend auf ihre Entdeckung warten. [90] Copyright (2014), mit Genehmigung der American Association for Advancement of Science
Toxizität von GFNs in Organen
Die Toxizität und Biokompatibilität von GFNs wurde durch theoretische und Tiermodellstudien beobachtet und bewertet. Gegenwärtig gibt es eine Fülle von Daten, die die Toxizität von GFNs in verschiedenen Organen oder Systemen bei Tieren belegen, so dass es schwierig ist, alle Daten in dieser Übersicht aufzulisten. Daher haben wir eine Reihe von Literatur zusammengefasst und einige toxikologische In-vivo-Studien von GFNs ausgewählt, die in Tabelle 1 aufgeführt sind.
Tabelle 1 Toxizität von GFNs in Organen
Tabelle in voller Größe
Toxizität in inneren Organen
GO kann durch Beeinträchtigung der normalen physiologischen Funktionen wichtiger Organe zu einer akuten Entzündungsreaktion und chronischen Verletzungen führen [32, 81]. Versuche mit oraler Sonde zeigten keine nachweisbare Aufnahme von GO durch den Magen-Darm-Trakt [95]. Interessanterweise verursachte eine niedrige GO-Dosis ernsthafte Schäden im Magen-Darm-Trakt, nachdem mütterliche Mäuse eine GO-Suspension anstelle einer hohen GO-Dosis getrunken hatten, da eine niedrige GO-Dosis ohne Agglomeration leicht an der Magen-Darm-Oberfläche haften und durch seine reichliche Menge Zerstörung verursachen kann scharfe Kanten [53]. GFNs verursachten eine Entzündung und verblieben am Tag 90 nach einer einmaligen intratrachealen Instillation in der Lunge und wurden sogar durch eine reine Naseninhalation in die Lungenlymphknoten transloziert [96, 97]. Eine hohe Dosis von Aggregaten bildendem GO kann pulmonale Blutgefäße blockieren und zu Atemnot führen [50, 98] und bei hohen Konzentrationen von 1 und 2 mg/kg KG wurden Thrombozytenthromben bei intravenöser Injektion beobachtet [89]. Berichten zufolge störte GO die Alveolar-Kapillar-Schranke, wodurch Entzündungszellen in die Lunge eindringen und die Freisetzung proinflammatorischer Zytokine stimulieren konnten [99]. Fibrose und Entzündungen konnten durch die erhöhten Spiegel der Proteinmarker Kollagen1, Gr1, CD68 und CD11b in der Lunge nachgewiesen werden. Die Verwendung von Tween 80 zum Dispergieren von FLG oder einem Pluronic-Surfactant zum Dispergieren von Graphen wurde vorgeschlagen, um die Wahrscheinlichkeit der Bildung von Lungenfibrose in Zellen oder Mäusen zu verringern, während Lungenfibrose beobachtet wurde, wenn Graphen mit Rinderserumalbumin (BSA) suspendiert wurde [100]. Darüber hinaus können radioaktive Isotope in die Lunge gelangen, begleitet von einer Tiefenverteilung von 125 I-NGO in der Lunge, die sich dort ablagern und zu Mutationen und Krebs führen können [30]. Neuere Veröffentlichungen behaupteten jedoch keine offensichtlichen pathologischen Veränderungen bei Mäusen, die niedrigen Dosen von GO und funktionalisiertem Graphen durch intravenöse Injektion ausgesetzt waren, einschließlich aminiertes GO (GO-NH2), Poly(acrylamid)-funktionalisiertes GO (GO-PAM), Poly(acrylsäure) )-funktionalisiertes GO (GO-PAA) und GO-PEG; nur GO-PEG und GO-PAA induzierten in vivo eine geringere Toxizität als reines GO [31, 79, 89]. Somit beeinflussen die funktionellen Gruppen von GFNs und die Arbeitskonzentration bzw. der Aggregatzustand maßgeblich die Toxizität von GFNs. In letzter Zeit werden die Möglichkeiten zur Modifizierung der funktionellen Gruppe von GFNs, zur Verringerung der Arbeitskonzentration oder zur Änderung des Aggregatzustands üblicherweise verwendet, um die Toxizität von GFNs zu verringern.
Toxizität im Zentralnervensystem
Graphen hat der Neurochirurgie durch die Anwendung der Wirkstoff-/Genabgabe zur Behandlung von Hirntumoren, intrakraniellen und spinalen biokompatiblen Geräten, Biosensorik und Bioimaging-Techniken großen Nutzen gebracht. Studien zu den Möglichkeiten oder Risiken von Graphen im Gehirn sind aufgetaucht. Im Hühnerembryomodell verringerten unberührte Graphenflocken den Ribonukleinsäurespiegel und die Geschwindigkeit der Desoxyribonukleinsäuresynthese, was zu schädlichen Auswirkungen auf die Entwicklung des Hirngewebes führte und die atypische Ultrastruktur im Gehirn beobachtet wurde [101]. Die neueren Forschungen zu GFNs im Zentralnervensystem befassen sich hauptsächlich mit der Anwendung und nicht mit der Toxizität. Die Daten der toxischen Studie zu GFNs sind im Gange.
Toxizität im Reproduktions- und Entwicklungssystem
Reines Graphen reduzierte die Vaskularisierung des Herzens und die Dichte verzweigter Gefäße nach Injektion in befruchtete Hühnereier und anschließender Inkubation für 19 d [101]. GO und rGO schädigen Zebrafischembryonen, indem sie die Embryoschlüpfrate und Körperlänge konzentrationsabhängig beeinflussen. Obwohl bei exponierten Zebrafischembryonen keine offensichtliche Missbildung oder Mortalität beobachtet wurde [102], haftete GO an und wurde in das Chorion der Zebrafischembryonen eingewickelt, was zu einer bemerkenswerten Hypoxie und Schlupfverzögerung führte. GO-Aggregate wurden in vielen Organellen wie Augen, Herz, Dottersack und Schwanz der Embryonen zurückgehalten, und in diesen Regionen wurden Apoptose und Bildung reaktiver Sauerstoffspezies (ROS) beobachtet [103].
Die GFNs üben unterschiedliche toxikologische Wirkungen auf das männliche oder weibliche Fortpflanzungssystem aus. Die Daten zeigten, dass GO selbst bei hoher Dosis durch intraabdominale Injektion nur sehr geringe oder fast keine toxischen Wirkungen auf die männliche Reproduktion ausübte [66]. Darüber hinaus veränderte rGO die Serumöstrogenspiegel von nicht schwangeren weiblichen Mäusen nicht. Bei der weiblichen Maus ist der Zustand anders: Mäusemuttertiere könnten nach rGO-Injektion vor der Paarung oder während der frühen Trächtigkeit gesunde Nachkommen gebären, und nur wenige abnormale Föten
Einfluss der Hämokompatibilität
Die Freisetzung von GO ins Blut ist unvermeidlich. Es wurde festgestellt, dass die Hämokompatibilität von GO von der funktionellen Beschichtung und den Expositionsbedingungen abhängt. GO mit einer Größe im Submikronbereich führte zu der größten hämolytischen Aktivität, während aggregiertes Graphen die geringste hämolytische Reaktion induzierte. Reines Graphen und GO zeigten eine hämolytische Wirkung bis zu 75 µg/ml [104]. GO-Polyethylenimin (GO-PEI) zeigte eine bemerkenswerte Toxizität durch Bindung an HSA, selbst bei 1,6 µg/mL [105]. Carboxyliertes Graphenoxid (GO-COOH) zeigte eine signifikante Zytotoxizität gegenüber T-Lymphozyten bei Konzentrationen über 50 µg/ml und eine gute Biokompatibilität unter 25 µg/ml, während GO-Chitosan die hämolytische Aktivität nahezu hemmte [106]. Das entsprechende Risiko der Hämokompatibilität war bisher weitgehend unbekannt.
Zusammenfassend lässt sich sagen, dass die durch GFNs induzierte Lungenschädigung in mehreren Studien untersucht wurde, deren Ergebnisse eine entzündliche Zellinfiltration, Lungenödeme und Granulombildung in der Lunge gezeigt haben. Allerdings wurden nur wenige spezifische Studien an anderen Organen wie Leber, Milz und Niere ausgewertet, und die Verletzungssymptome, der Schadensindex und das Ausmaß der Schädigung dieser inneren Organe wurden nicht vollständig untersucht. Darüber hinaus sind Studien zur Neurotoxizität von GFNs recht selten; keine Daten haben ergeben, welche Nerven oder Hirnareale geschädigt sind, noch wurden die entsprechenden Verhaltensmanifestationen untersucht. Die Entwicklungstoxizität von GFNs kann zu strukturellen Anomalien, Wachstumsverzögerungen, Verhaltens- und Funktionsanomalien und sogar zum Tod führen. Eine Studie zur Reproduktions- und Entwicklungstoxizität von GFNs wird von großer Bedeutung sein und in Zukunft große Aufmerksamkeit erhalten. Fast alle Toxizitätsstudien von GFNs waren Kurzzeitexperimente, und keine Studien haben chronische toxische Langzeitschäden untersucht. Basierend auf Studien zur Toxizität anderer Nanomaterialien kann die langfristige Exposition mit GFNs jedoch ein wichtiger gesundheitsschädlicher Faktor sein [107–109]. Daher ist die Langzeitstudie von GFNs notwendig.
Toxizität von GFNs in Zellmodellen
Die Zytotoxizität von GFNs in vitro wurde in verschiedenen Zellen nachgewiesen, um die Lebensfähigkeit und Morphologie der Zellen zu verändern, die Membranintegrität zu zerstören und DNA-Schäden zu induzieren [110–112]. GO oder rGO verringern die Zelladhäsion; Zellapoptose induzieren; und dringen in Lysosomen, Mitochondrien, Zellkerne und Endoplasma ein [113]. GQDs traten in Zellen ein und induzierten DNA-Schäden durch die erhöhte Expression von p53-, Rad-51- und OGG1-Proteinen in NIH-3-T3-Zellen [87]. GQDs wiesen jedoch keine signifikante Toxizität gegenüber menschlichen Brustkrebszelllinien (bei einer Dosis von 50 µg/ml) oder menschlichen neuralen Stammzellen (bei einer Dosis von 250 µg/ml) auf [114, 115]. GO-Derivate verringerten die Expression differenzieller Gene, die für die Struktur und Funktion der Zellmembran verantwortlich sind, wie die Regulierung des Aktinzytoskeletts, die fokale Adhäsion und die Endozytose, dramatisch [89]. In Phäochromozytomzellen der Ratte (PC12-Zellen) verursachten Graphen und rGO zytotoxische Effekte und mitochondriale Schäden, wie die Freisetzung von Lactatdehydrogenase (LDH), eine erhöhte Aktivierung von Caspase-3 und die Bildung von ROS [82, 116] .
Graphen kann die Lebensfähigkeit der Zellen erhöhen [117] oder den Zelltod verursachen [118], abhängig von der Zelllinie, der Art des Graphenmaterials und der Dosierung. GO-Zytotoxizität wurde in humanen Fibroblasten und Lungenepithelzellen bei Konzentrationen über 20 µg/ml nach 24 h beobachtet, jedoch wurde eine minimale Toxizität bei A549-Zellen bei Konzentrationen über 50 µg/ml gefunden [119]. Die durch GO induzierten biologischen Reaktionen wie ROS, Malondialdehyd (MDA) und LDH nahmen zu, während Superoxiddismutase (SOD) in HeLa-Zellen dosisabhängig abnahm [120]. GO-molecular beacon (GO-MB) zeigte jedoch selbst bei 20 µg/ml in HeLa-Zellen eine geringe Zytotoxizität [121]. GO verringerte die Lebensfähigkeit von A549-Zellen, während die gleiche Konzentration und Expositionszeit die Zelllebensfähigkeit von CaCo2-Kolorektalkarzinomzellen erhöhte [122]. Eine andere Studie berichtete, dass GO die Differenzierung von SH-SY5Y dramatisch verbesserte, begleitet von einer Erhöhung der Neuritenlänge und der Expression des neuronalen Markers MAP2 bei niedrigen Konzentrationen, dass GO jedoch die Lebensfähigkeit von SH-SY5Y-Zellen bei hohen Dosen (≥80 mg/ml) unterdrückte. [123]. Funktionalisierte Beschichtungen auf GO, wie GO-PEG [124] und GO-Chitosan [125], können die Zytotoxizität der Partikel durch Hemmung der Wechselwirkungen zwischen Zellen stark abschwächen.
Die Toxizität von GFNs in vitro ist in Tabelle 2 zusammengefasst. Die Daten zur Zytotoxizität von Graphen-Nanomaterialien sind gegensätzlich und unterschiedliche Eigenschaften beeinflussen die Ergebnisse. Die Mechanismen und Einflussfaktoren der Toxizität müssen im Detail aufgeklärt werden.
Ursprünge der GFN-Toxizität
Berichten zufolge beeinflussen die Eigenschaften von Graphen, einschließlich seiner Konzentration, lateralen Dimension, Oberflächenstruktur, funktionellen Gruppen, Reinheit und Proteinkorona, stark seine Toxizität in biologischen Systemen [2, 7, 104, 126–129].
Konzentration
Zahlreiche Ergebnisse haben gezeigt, dass Graphenmaterialien bei Tieren und Zellen eine dosisabhängige Toxizität verursachen, wie Leber- und Nierenschäden, Lungengranulombildung, verminderte Zelllebensfähigkeit und Zellapoptose [130–134]. In-vivo-Studien zeigte GO keine offensichtliche Toxizität bei Mäusen, die einer niedrigen Dosis (0,1 mg) und einer mittleren Dosis (0,25 mg) ausgesetzt waren, induzierte jedoch bei einer hohen Dosis (0,4 mg) eine chronische Toxizität. Der hohe Gehalt an GO lagerte sich hauptsächlich in Lunge, Leber, Milz und Niere ab und war durch eine einzelne Schwanzveneninjektion von den Nieren nur schwer zu reinigen [135]. Interessanterweise führte eine Erhöhung der Dosis zu einer dramatischen Abnahme der hepatischen Aufnahme, jedoch zu einer Zunahme der pulmonalen Aufnahme von s-GO durch intravenöse Injektion [31], da die hohe Dosis von GO möglicherweise die Aufnahmesättigung übertraf oder die Masse an Plasma-Opsoninen verringerte , die folglich die hepatische Aufnahme unterdrückt. Darüber hinaus wurde in einer In-vitro-Studie berichtet, dass 20 µg/mL GO-Nanosheets innerhalb von 2 h nach Inkubation keine Zytotoxizität in A549 zeigten, eine höhere Konzentration (85 µg/mL) jedoch die Zellviabilität innerhalb von 24 h auf 50 % verringerte [136, 137] . Lüet al. zeigten auch, dass GO bei niedrigen Konzentrationen 96 h lang keine offensichtliche Zytotoxizität in einer humanen Neuroblastom-SH-SY5Y-Zelllinie aufwies, jedoch die Lebensfähigkeit der Zellen nach Behandlung mit 100 mg/ml GO für 96 h Inkubation stark auf 20 % abnahm [123] . Die Ergebnisse in HeLa-Zellen, NIH-3 T3-Zellen und Brustkrebszellen (SKBR3, MCF7), die mit Graphen-Nanobändern behandelt wurden, zeigten ebenfalls eine dosis- (10–400 mg/ml) und zeitabhängige (12–48 h) Abnahme der Zelllebensfähigkeit [138]. Zunehmende Konzentrationen von GO gelangten in die Lysosomen, Mitochondrien, das Endoplasma und den Zellkern [119]. Mehrere Daten deuteten darauf hin, dass rGO bei einer niedrigeren Dosis und zu einem frühen Zeitpunkt den Apoptose-vermittelten Zelltod verursachte, dass jedoch mit zunehmender Zeit/Dosis Nekrose vorherrschte [110, 135].
Seitenmaß
Nanopartikel mit einer Größe < 100 nm können in die Zelle gelangen, < 40 nm in den Zellkern und kleiner als < 35 nm können die Blut-Hirn-Schranke passieren [85]. Eine Studie zeigte, dass GO (588, 556, 148 nm) nicht in A549-Zellen eindrang und keine offensichtliche Zytotoxizität aufwies [112]. Wenn der Durchmesser von Graphen zwischen 100 ~ 500 nm liegt, kann die kleinste Größe die stärkste Toxizität verursachen, und wenn der Durchmesser unter 40 nm liegt, können die kleinsten Größen die sichersten sein. Beispielsweise könnte rGO mit einem Durchmesser von 11 ± 4 nm in den Kern der hMSCs eindringen und bei sehr niedrigen Konzentrationen von 0,1 und 1,0 mg/mL in 1 h Chromosomenaberrationen und DNA-Fragmentierung verursachen. rGO-Faltblätter mit Durchmessern von 3,8 ± 0,4 nm zeigten jedoch selbst bei einer hohen Dosis von 100 mg/ml nach 24 h keine nennenswerte Genotoxizität in hMSCs [118].
In einer In-vivo-Studie akkumulierte s-GO (100–500 nm) bevorzugt in der Leber, wohingegen l-GO (1–5 µm) hauptsächlich in der Lunge lokalisiert war, da l-GO größere GO-Proteinkomplexe bildete, die gefiltert wurden durch die pulmonalen Kapillargefäße nach intravenöser Injektion [31]. Angesichts der relativen lateralen Größen (205,8 nm, 146,8 nm und 33,78 nm) der drei GO-Nanoblätter bei gleicher Konzentration erfährt kleineres GO in Hela-Zellen eine viel größere Aufnahme als größeres GO [139]. Die hohe Aufnahme von s-GO veränderte sich in der Mikroumgebung der Zellen und induzierte folglich den größten Verlust der Lebensfähigkeit und den schwerwiegendsten oxidativen Stress unter drei Größen von GO-Proben [119]. Als Ergebnis zeigte eine Studie, dass GO größenabhängig die M1-Polarisierung von Makrophagen und proinflammatorische Reaktionen in vitro und in vivo induzierte. Größeres GO zeigte eine stärkere Adsorption an die Plasmamembran mit weniger Phagozytose, löste robuste Interaktionen mit TLRs aus und aktivierte NF-κB-Wege im Vergleich zu kleineren GO-Faltblättern, die eher von Zellen aufgenommen wurden [94]. Um den detaillierten Mechanismus, der diesen Effekten zugrunde liegt, aufzudecken, sind weitere Studien erforderlich, um den entscheidenden Mechanismus der lateralen Größe von Graphenmaterialien zu veranschaulichen.
Ursprünge der GFN-Toxizität
Berichten zufolge beeinflussen die Eigenschaften von Graphen, einschließlich seiner Konzentration, lateralen Dimension, Oberflächenstruktur, funktionellen Gruppen, Reinheit und Proteinkorona, stark seine Toxizität in biologischen Systemen [2, 7, 104, 126–129].
Konzentration
Zahlreiche Ergebnisse haben gezeigt, dass Graphenmaterialien bei Tieren und Zellen eine dosisabhängige Toxizität verursachen, wie Leber- und Nierenschäden, Lungengranulombildung, verminderte Zelllebensfähigkeit und Zellapoptose [130–134]. In-vivo-Studien zeigte GO keine offensichtliche Toxizität bei Mäusen, die einer niedrigen Dosis (0,1 mg) und einer mittleren Dosis (0,25 mg) ausgesetzt waren, induzierte jedoch bei einer hohen Dosis (0,4 mg) eine chronische Toxizität. Der hohe Gehalt an GO lagerte sich hauptsächlich in Lunge, Leber, Milz und Niere ab und war durch eine einzelne Schwanzveneninjektion von den Nieren nur schwer zu reinigen [135]. Interessanterweise führte eine Erhöhung der Dosis zu einer dramatischen Abnahme der hepatischen Aufnahme, jedoch zu einer Zunahme der pulmonalen Aufnahme von s-GO durch intravenöse Injektion [31], da die hohe Dosis von GO möglicherweise die Aufnahmesättigung übertraf oder die Masse an Plasma-Opsoninen verringerte , die folglich die hepatische Aufnahme unterdrückt. Darüber hinaus wurde in einer In-vitro-Studie berichtet, dass 20 µg/mL GO-Nanosheets innerhalb von 2 h nach Inkubation keine Zytotoxizität in A549 zeigten, eine höhere Konzentration (85 µg/mL) jedoch die Zellviabilität innerhalb von 24 h auf 50 % verringerte [136, 137] . Lüet al. zeigten auch, dass GO bei niedrigen Konzentrationen 96 h lang keine offensichtliche Zytotoxizität in einer humanen Neuroblastom-SH-SY5Y-Zelllinie aufwies, jedoch die Lebensfähigkeit der Zellen nach Behandlung mit 100 mg/ml GO für 96 h Inkubation stark auf 20 % abnahm [123] . Die Ergebnisse in HeLa-Zellen, NIH-3 T3-Zellen und Brustkrebszellen (SKBR3, MCF7), die mit Graphen-Nanobändern behandelt wurden, zeigten ebenfalls eine dosis- (10–400 mg/ml) und zeitabhängige (12–48 h) Abnahme der Zelllebensfähigkeit [138]. Zunehmende Konzentrationen von GO gelangten in die Lysosomen, Mitochondrien, das Endoplasma und den Zellkern [119]. Mehrere Daten deuteten darauf hin, dass rGO bei einer niedrigeren Dosis und zu einem frühen Zeitpunkt den Apoptose-vermittelten Zelltod verursachte, dass jedoch mit zunehmender Zeit/Dosis Nekrose vorherrschte [110, 135].
Seitenmaß
Nanopartikel mit einer Größe < 100 nm können in die Zelle gelangen, < 40 nm in den Zellkern und kleiner als < 35 nm können die Blut-Hirn-Schranke passieren [85]. Eine Studie zeigte, dass GO (588, 556, 148 nm) nicht in A549-Zellen eindrang und keine offensichtliche Zytotoxizität aufwies [112]. Wenn der Durchmesser von Graphen zwischen 100 ~ 500 nm liegt, kann die kleinste Größe die stärkste Toxizität verursachen, und wenn der Durchmesser unter 40 nm liegt, können die kleinsten Größen die sichersten sein. Beispielsweise könnte rGO mit einem Durchmesser von 11 ± 4 nm in den Kern der hMSCs eindringen und bei sehr niedrigen Konzentrationen von 0,1 und 1,0 mg/mL in 1 h Chromosomenaberrationen und DNA-Fragmentierung verursachen. rGO-Faltblätter mit Durchmessern von 3,8 ± 0,4 nm zeigten jedoch selbst bei einer hohen Dosis von 100 mg/ml nach 24 h keine nennenswerte Genotoxizität in hMSCs [118].
In einer In-vivo-Studie akkumulierte s-GO (100–500 nm) bevorzugt in der Leber, wohingegen l-GO (1–5 µm) hauptsächlich in der Lunge lokalisiert war, da l-GO größere GO-Proteinkomplexe bildete, die gefiltert wurden durch die pulmonalen Kapillargefäße nach intravenöser Injektion [31]. Angesichts der relativen lateralen Größen (205,8 nm, 146,8 nm und 33,78 nm) der drei GO-Nanoblätter bei gleicher Konzentration erfährt kleineres GO in Hela-Zellen eine viel größere Aufnahme als größeres GO [139]. Die hohe Aufnahme von s-GO veränderte sich in der Mikroumgebung der Zellen und induzierte folglich den größten Verlust der Lebensfähigkeit und den schwerwiegendsten oxidativen Stress unter drei Größen von GO-Proben [119]. Als Ergebnis zeigte eine Studie, dass GO größenabhängig die M1-Polarisierung von Makrophagen und proinflammatorische Reaktionen in vitro und in vivo induzierte. Größeres GO zeigte eine stärkere Adsorption an die Plasmamembran mit weniger Phagozytose, löste robuste Interaktionen mit TLRs aus und aktivierte NF-κB-Wege im Vergleich zu kleineren GO-Faltblättern, die eher von Zellen aufgenommen wurden [94]. Um den detaillierten Mechanismus, der diesen Effekten zugrunde liegt, aufzudecken, sind weitere Studien erforderlich, um den entscheidenden Mechanismus der lateralen Größe von Graphenmaterialien zu veranschaulichen.
Ursprünge der GFN-Toxizität
Berichten zufolge beeinflussen die Eigenschaften von Graphen, einschließlich seiner Konzentration, lateralen Dimension, Oberflächenstruktur, funktionellen Gruppen, Reinheit und Proteinkorona, stark seine Toxizität in biologischen Systemen [2, 7, 104, 126–129].
Konzentration
Zahlreiche Ergebnisse haben gezeigt, dass Graphenmaterialien bei Tieren und Zellen eine dosisabhängige Toxizität verursachen, wie Leber- und Nierenschäden, Lungengranulombildung, verminderte Zelllebensfähigkeit und Zellapoptose [130–134]. In-vivo-Studien zeigte GO keine offensichtliche Toxizität bei Mäusen, die einer niedrigen Dosis (0,1 mg) und einer mittleren Dosis (0,25 mg) ausgesetzt waren, induzierte jedoch bei einer hohen Dosis (0,4 mg) eine chronische Toxizität. Der hohe Gehalt an GO lagerte sich hauptsächlich in Lunge, Leber, Milz und Niere ab und war durch eine einzelne Schwanzveneninjektion von den Nieren nur schwer zu reinigen [135]. Interessanterweise führte eine Erhöhung der Dosis zu einer dramatischen Abnahme der hepatischen Aufnahme, jedoch zu einer Zunahme der pulmonalen Aufnahme von s-GO durch intravenöse Injektion [31], da die hohe Dosis von GO möglicherweise die Aufnahmesättigung übertraf oder die Masse an Plasma-Opsoninen verringerte , die folglich die hepatische Aufnahme unterdrückt. Darüber hinaus wurde in einer In-vitro-Studie berichtet, dass 20 µg/mL GO-Nanosheets innerhalb von 2 h nach Inkubation keine Zytotoxizität in A549 zeigten, eine höhere Konzentration (85 µg/mL) jedoch die Zellviabilität innerhalb von 24 h auf 50 % verringerte [136, 137] . Lüet al. zeigten auch, dass GO bei niedrigen Konzentrationen 96 h lang keine offensichtliche Zytotoxizität in einer humanen Neuroblastom-SH-SY5Y-Zelllinie aufwies, jedoch die Lebensfähigkeit der Zellen nach Behandlung mit 100 mg/ml GO für 96 h Inkubation stark auf 20 % abnahm [123] . Die Ergebnisse in HeLa-Zellen, NIH-3 T3-Zellen und Brustkrebszellen (SKBR3, MCF7), die mit Graphen-Nanobändern behandelt wurden, zeigten ebenfalls eine dosis- (10–400 mg/ml) und zeitabhängige (12–48 h) Abnahme der Zelllebensfähigkeit [138]. Zunehmende Konzentrationen von GO gelangten in die Lysosomen, Mitochondrien, das Endoplasma und den Zellkern [119]. Mehrere Daten deuteten darauf hin, dass rGO bei einer niedrigeren Dosis und zu einem frühen Zeitpunkt den Apoptose-vermittelten Zelltod verursachte, dass jedoch mit zunehmender Zeit/Dosis Nekrose vorherrschte [110, 135].
Seitenmaß
Nanopartikel mit einer Größe < 100 nm können in die Zelle gelangen, < 40 nm in den Zellkern und kleiner als < 35 nm können die Blut-Hirn-Schranke passieren [85]. Eine Studie zeigte, dass GO (588, 556, 148 nm) nicht in A549-Zellen eindrang und keine offensichtliche Zytotoxizität aufwies [112]. Wenn der Durchmesser von Graphen zwischen 100 ~ 500 nm liegt, kann die kleinste Größe die stärkste Toxizität verursachen, und wenn der Durchmesser unter 40 nm liegt, können die kleinsten Größen die sichersten sein. Beispielsweise könnte rGO mit einem Durchmesser von 11 ± 4 nm in den Kern der hMSCs eindringen und bei sehr niedrigen Konzentrationen von 0,1 und 1,0 mg/mL in 1 h Chromosomenaberrationen und DNA-Fragmentierung verursachen. rGO-Faltblätter mit Durchmessern von 3,8 ± 0,4 nm zeigten jedoch selbst bei einer hohen Dosis von 100 mg/ml nach 24 h keine nennenswerte Genotoxizität in hMSCs [118].
In einer In-vivo-Studie akkumulierte s-GO (100–500 nm) bevorzugt in der Leber, wohingegen l-GO (1–5 µm) hauptsächlich in der Lunge lokalisiert war, da l-GO größere GO-Proteinkomplexe bildete, die gefiltert wurden durch die pulmonalen Kapillargefäße nach intravenöser Injektion [31]. Angesichts der relativen lateralen Größen (205,8 nm, 146,8 nm und 33,78 nm) der drei GO-Nanoblätter bei gleicher Konzentration erfährt kleineres GO in Hela-Zellen eine viel größere Aufnahme als größeres GO [139]. Die hohe Aufnahme von s-GO veränderte sich in der Mikroumgebung der Zellen und induzierte folglich den größten Verlust der Lebensfähigkeit und den schwerwiegendsten oxidativen Stress unter drei Größen von GO-Proben [119]. Als Ergebnis zeigte eine Studie, dass GO größenabhängig die M1-Polarisierung von Makrophagen und proinflammatorische Reaktionen in vitro und in vivo induzierte. Größeres GO zeigte eine stärkere Adsorption an die Plasmamembran mit weniger Phagozytose, löste robuste Interaktionen mit TLRs aus und aktivierte NF-κB-Wege im Vergleich zu kleineren GO-Faltblättern, die eher von Zellen aufgenommen wurden [94]. Um den detaillierten Mechanismus, der diesen Effekten zugrunde liegt, aufzudecken, sind weitere Studien erforderlich, um den entscheidenden Mechanismus der lateralen Größe von Graphenmaterialien zu veranschaulichen.
Oberflächenstruktur
GFNs besitzen sehr unterschiedliche Oberflächenchemien. Zum Beispiel ist die unberührte Graphenoberfläche hydrophob, die GO-Oberfläche ist mit Carboxylatgruppen teilweise hydrophob [140–142] und rGO hat eine mittlere Hydrophilie [143]. Es wurde beobachtet, dass GFNs die Funktion und Struktur von Zellmembranen und Proteinen wahrscheinlich durch außergewöhnlich starke molekulare Interaktionen mit Zellen stören [2, 91]. Beispielsweise bindet rGO an Zellmembranen, stimuliert Rezeptoren und aktiviert mitochondriale Signalwege, wodurch Apoptose induziert wird [110, 111, 144]. Begrenzte Beweise zeigten, dass GO aufgrund des hohen Sauerstoffgehalts, der glatteren Kanten und der hydrophilen Eigenschaften der ersteren Spezies kleiner und weniger toxisch als rGO ist [104, 145, 146]. Aufgrund der unterschiedlichen Oberflächenoxidationsstufen von GO und rGO könnte GO, das eine unterschiedliche Hydrophilie besitzt, leicht von HepG2-Zellen internalisiert und aufgenommen werden. Im Gegensatz dazu konnte rGO mit offensichtlicher Hydrophobie ohne (oder mit geringerer) Aufnahme an Zelloberflächen adsorbiert und aggregiert werden [110]. Aufgrund der starken π-π-Stapelwechselwirkungen ist Graphen in der Lage, viele Reste des Proteins zu brechen, insbesondere die aromatischen, wie das Villin-Kopfstück (HP), F10, W23 und F35. Die Sekundär- und Tertiärstrukturen des Proteins liegen größtenteils auf der Graphenoberfläche und stören die Struktur und Funktion des Proteins [41] (Abb. 2). Außerdem kann GO zwischen die Basenpaare doppelsträngiger DNA inserieren und den genetischen Informationsfluss auf molekularer Ebene stören, was eine der Hauptursachen für die mutagene Wirkung von GO sein könnte [7, 112, 146, 147] .
Abb. 2
Figur 2
Eine repräsentative Trajektorie von HP35, die an Graphen adsorbiert. (a) Repräsentative Momentaufnahmen zu verschiedenen Zeitpunkten. Die Proteine sind in Cartoons mit roter Helix und grüner Schleife dargestellt, und das Graphen ist in Weizen dargestellt. Die aromatischen Reste, die die π-π-Stapelwechselwirkungen bilden, sind in Blau dargestellt, andere in Grün. (b) Die Kontaktfläche von HP35 mit dem Graphen. (c) Die RMSD von HP35 aus seiner nativen Struktur und der Anzahl der Reste in der α-Helix-Struktur. Hier werden die Sekundärstrukturen vom DSSP-Programm bestimmt. (d) Der Abstand zwischen dem Graphen und den aromatischen Resten, einschließlich F35, W23, F10, F17 und F06. Um den Adsorptionsprozess klarer darzustellen, wurde die χ-Achse abgeschnitten und neu skaliert. [41] Copyright (2011), mit Genehmigung von Journal of Physical of Chemistry
Bild in voller Größe
Aufladen
Eine Reihe von Studien haben die Bedeutung der GO-Oberflächenladung aufgrund ihrer Fähigkeit, den Internalisierungs- und Aufnahmemechanismus von Zellen zu beeinflussen, hervorgehoben [148–150]. Die Internalisierung von GO war in Nicht-Phagozyten vernachlässigbar, was wahrscheinlich auf die starke elektrostatische Abstoßung zwischen dem negativ geladenen GO und der Zelloberfläche zurückzuführen war [34]. Andere haben jedoch vorgeschlagen, dass negativ geladene Nanopartikel in nicht-phagozytische Zellen internalisiert werden können, indem sie an verfügbare kationische Stellen auf der Zelloberfläche binden und von Scavenger-Rezeptoren aufgenommen werden [110, 146, 150]. Berichten zufolge verursachen GO/GS-Partikel morphologische Veränderungen und eine signifikante Lyse, was zu einer hohen Hämolyse in roten Blutkörperchen (RBCs) führt. Die Zerstörung der RBC-Membran wird wahrscheinlich auf die starken elektrostatischen Wechselwirkungen zwischen den negativ geladenen Sauerstoffgruppen auf der GO/GS-Oberfläche und positiv geladenen Phosphatidylcholin-Lipiden auf der RBC-Außenmembran zurückgeführt [106].
Funktionalisierung
Studien bestätigten, dass die Funktionalisierung mit PEG [52], PEGyliertem Poly-L-Lysin (PLL) [151], Poly(ε-caprolacton) [152], Polyvinylalkohol [3], Pluronic [153], Amin [98], Carboxyl , und Dextran [79]-Gruppen verringert die Toxizität stark und verbessert die Biokompatibilität von Graphen. In-vivo-Ergebnisse zeigten, dass nach der subkutanen Injektion von GO-Pluronic Hydrogel nur eine leichte chronische Entzündung auftrat und nach der intravenösen Injektion von GO-DEX keine merkliche Kurzzeittoxizität getestet wurde [79, 154]. PEGyliertes GS induzierte keine nennenswerte Toxizität bei Mäusen, die 3 Monate lang gegen 20 mg/kg exponiert wurden, wie durch Blutbiochemie und histologische Untersuchungen bewertet wurde, und zeigte eine relativ geringe Retention im RES [52, 155]. Die Beschichtung von GO mit Chitosan eliminierte fast die hämolytische Aktivität im Blut [39]. Darüber hinaus linderte die PEG-Beschichtung wirksam GO-induzierte akute Gewebeverletzungen; verminderte GO-Aggregation und -Retention in Leber, Lunge und Milz; und förderte die Clearance von GO [81], GO-DEX [79] und fluoriertem Graphenoxid (FGO) [156].
In vitro zeigten mehrere Zellfunktionsassays klare Hinweise darauf, dass die Oberflächenfunktionalisierung von reinem Graphen oder GO entscheidend für die Reduzierung der starken Toxizitätseffekte war [91]. PEG-GO, PEI-GO und LA-PEG-GO schädigten menschliche Lungenfibroblastenzellen weniger als GO [148]. PEG-GO zeigte keine Zytotoxizität gegenüber mehreren Zellkulturen wie Glioblastomzellen (U87MG), Brustkrebszellen (MCF-7), menschlichen Ovarialkarzinomzellen (OVCAR-3), Dickdarmkrebszellen (HCT-116) und lymphoblastoiden Zellen (RAJI), in Konzentrationen bis 100 µg/ml [119, 157, 158]. GQDs-PEG zeigte selbst bei sehr hohen Konzentrationen (200 µg/ml) eine sehr geringe oder keine Toxizität gegenüber Lungen- und Gebärmutterhalskrebszellen [159]. Da es sich jedoch um ein nicht biologisch abbaubares Material mit großem Potenzial zur zellulären Internalisierung handelt, sind weitere Untersuchungen erforderlich, um die möglichen langfristigen Nebenwirkungen von funktionalisiertem Graphen zu bewerten.
Aggregationen und Sedimentation
Berichten zufolge neigen Nanomaterialien dazu, eher Aggregate als einzelne Einheiten zu bilden, insbesondere unter physiologischen Bedingungen. GS-Oberflächen ließen im Vergleich zu GO weniger Erythrozyten anheften, und GS hatte die geringere hämolytische Aktivität für die Bildung von mehr wässrigen Aggregaten. Im Gegensatz dazu hemmte die schnelle Sedimentation und Aggregatbildung von GS die Nährstoffverfügbarkeit von menschlichen Hautfibroblastenzellen, die auf dem Boden von Wells gezüchtet wurden [106]. Daher üben die Aggregationen und Sedimentation von Graphenpartikeln unterschiedliche Wirkungen auf verschiedene Zellen aus.
Verunreinigungen
Die Reinheit des Nanomaterials ist ein wichtiger Aspekt, da verbleibende kontaminierende Metalle für die beobachtete Toxizität verantwortlich sein können und nicht das Nanomaterial selbst, was zu widersprüchlichen Daten zur Zytotoxizität von GFNs geführt hat [35, 160]. Herkömmlich hergestelltes GO enthält oft hohe Mengen an Mn2+ und Fe2+, die für Zellen stark mutagen sind. Die unspezifische Freisetzung dieser Ionen aus traditionell hergestelltem GO könnte zu ungewöhnlich hohen Zytotoxizitäten und DNA-Frakturen führen [39]. Insbesondere Peng et al. [161] produzierten hochreines GO mit nur 0.025 ppm Mn2+ und 0.13 ppm Fe2+, und Hanene et al. [162] erfanden eine neue Methode zur Herstellung hochreiner einschichtiger GO-Schichten mit guter wässriger Dispergierbarkeit und kolloidaler Stabilität. Mit diesen neuen Methoden hergestelltes GO induzierte in vitro keine signifikanten zytotoxischen Reaktionen (bei Expositionsdosen von bis zu 100 µg/ml), und in vivo wurde keine offensichtliche Entzündungsreaktion oder Granulombildung (Expositionsdosen bis zu 50 µg/Tier) beobachtet. Daher verdient die Reinheit von GFNs Aufmerksamkeit und ist ein wichtiger Schritt zur Bestimmung von GFNs, die an Bioanwendungen beteiligt sind.
Protein-Corona-Effekt
Aufgrund der hohen freien Oberflächenladung können Nanomaterialien mit Proteinen in biologischen Systemen leicht „Coronas“ bilden [163, 164]. Es wird vermutet, dass die Proteinkorona die Zirkulation, Verteilung, Clearance und Toxizität von Nanopartikeln beeinflusst. Mehrere Veröffentlichungen berichteten, dass GO mit adsorbierten Plasmaproteinen im Serum GO-Protein-Coronas bildet und diese GO-Protein-Coronas eine wichtige Rolle bei der Entscheidung über das Schicksal des biokinetischen Verhaltens von GO in vivo spielen. Solche GO-Protein-Coronas können die Adhäsion von GO an Endothel- und Immunzellen sowohl durch spezifische als auch durch unspezifische Wechselwirkungen regulieren [165]. Grundsätzlich helfen Immunglobulin G und Komplementproteine in der Proteinkorona, Nanopartikel in Immunzellen zu reorganisieren, wodurch die Partikel vom RES verschlungen werden, und IgG-beschichtetes GO wurde entweder durch spezifische oder unspezifische Interaktionen mit Zellmembranrezeptoren aufgenommen [31, 165]. Eine andere Studie ergab jedoch, dass GO nicht direkt im Darmtrakt an Schleimhautepithelzellen anhaften konnte, nachdem die Filialmäuse eine wässrige GO-Lösung getrunken hatten, da reichlich Proteine in der Milch an der Oberfläche des GO adsorbiert waren und somit deren direkte Wechselwirkung mit dem Schleimhautepithelzellen [53]. Proteinkorona milderte die Zytotoxizität von GO, indem sie seine physikalische Wechselwirkung mit der Zellmembran begrenzte und die zellmorphologische Schädigung in HeLa-, THP-1- und A549-Zellen reduzierte [166–168]. Die zytotoxische Wirkung wurde stark reduziert, wenn GO mit FBS vorbeschichtet und mit Zellen inkubiert wurde; Fast ∼ 90 % Überleben wurde mit 100 µg/ml FBS-beschichtetem GO und 100 % Überleben mit 20 µg/ml FBS-beschichtetem GO beobachtet. Ähnliche Trends wurden für GO beobachtet, das von BSA abgedeckt wird [166, 169]. Konsequenterweise könnte zusätzliches Serum die Toxizität von reinem GO in J774.A1-Zellen in einer Dosis von 4 µg/ml neutralisieren, was zu einer Abnahme der Zellzahl um 52,5 % im Vergleich zu unbehandelten Zellen führte [89].
Nach Durchsicht vieler Studien kann der Schluss gezogen werden, dass die Toxizität von Graphen von mehreren Faktoren beeinflusst wird. Diese Faktoren kombinierten die Toxizität von GFNs in vielen Fällen stark. Wissenschaftliche Studien erfordern oft die eindeutige Identifizierung von Ursache und Wirkung, wobei jeweils nur ein Faktor unterschiedlich sein sollte, damit die Wirkung dieses einzelnen Faktors bestimmt werden kann. In einigen Veröffentlichungen wurden jedoch mehrere Faktoren, die die Toxizität von GFNs beeinflussen, gleichzeitig untersucht, was zu verwirrenden Ergebnissen führte.
Mögliche Toxizitätsmechanismen von GFNs
Obwohl einige physikalisch-chemische Eigenschaften und die Toxizität von GFNs von vielen Wissenschaftlern gut untersucht wurden, bleiben die genauen Mechanismen, die der Toxizität von GFNs zugrunde liegen, unklar. Eine schematische Darstellung der Hauptmechanismen der Zytotoxizität von GFNs ist in Abb. 3 dargestellt.
Physische Zerstörung
Graphen ist im Vergleich zu anderen kugelförmigen oder eindimensionalen Nanopartikeln aufgrund seiner zweidimensionalen Struktur mit sp2-Kohlenstoffen ein einzigartiges Nanomaterial. Die physikalische Wechselwirkung von Graphen-Nanopartikeln mit Zellmembranen ist eine der Hauptursachen für die Zytotoxizität von Graphen [7, 170, 171]. Graphen hat aufgrund seiner günstigen Oberflächenkrümmung eine hohe Fähigkeit, an die α-helikalen Strukturen von Peptiden zu binden [172]. Bei Konzentrationen über 75 µg/ml haftete reines Graphen weitgehend an der Oberfläche von RAW 264.7-Zellen und führte zu einer abnormalen Dehnung der Zellmembran [104]. Die starken hydrophoben Wechselwirkungen von GFNs mit der Zellmembran führen zur morphologischen Erweiterung der filopodialen und zytoskelettalen F-Aktin-Dysfunktion. Darüber hinaus können die scharfen Kanten von GNS als „Klingen“ fungieren, die bakterielle Zellmembranen einführen und durchschneiden [173]. Darüber hinaus schädigte GO auch direkt die äußere Membran von E. coli-Bakterien, was zur Freisetzung intrazellulärer Komponenten führte [173]. Die TEM-Bildgebung zeigte jedoch, dass die Vorbeschichtung von GO mit FBS die Zerstörung von Zellmembranen beseitigte [166].
ROS-Produktion führt zu oxidativem Stress
Oxidativer Stress entsteht, wenn steigende ROS-Spiegel die Aktivität antioxidativer Enzyme wie Katalase, SOD oder Glutathionperoxidase (GSH-PX) überfordern [174]. ROS wirken als Second Messenger in vielen intrazellulären Signalkaskaden und führen zu zellulären makromolekularen Schäden wie Membranlipidabbau, DNA-Fragmentierung, Proteindenaturierung und mitochondrialer Dysfunktion, die den Zellstoffwechsel und die Signalübertragung stark beeinflussen [175–177]. Die Interaktionen von GO mit Zellen können zu einer übermäßigen ROS-Erzeugung führen, die den ersten Schritt in den Mechanismen der Karzinogenese, Alterung und Mutagenese darstellt [83, 122]. Oxidativer Stress spielte eine signifikante Rolle bei der GO-induzierten akuten Lungenschädigung [30], und die durch oxidativen Stress verursachten Entzündungsreaktionen traten oft bei Exposition gegenüber GFNs auf [133, 177, 178]. Die Aktivität von SOD und GSH-PX nahm nach Exposition gegenüber GO zeit- und dosisabhängig ab [82, 106, 119]. Ebenso war oxidativer Stress die Hauptursache für Apoptose und DNA-Schäden, nachdem HLF-Zellen GO ausgesetzt wurden [148]. Sowohl die Mitogen-aktivierte Proteinkinase (MAPK) (JNK, ERK und p38) als auch die TGF-beta-bezogenen Signalwege wurden durch die ROS-Erzeugung in reinen Graphen-behandelten Zellen ausgelöst, begleitet von der Aktivierung von Bim und Bax, zwei Pro -apoptotische Mitglieder der Bcl-2-Proteinfamilie. Dadurch wurden Caspase-3 und seine nachgeschalteten Effektorproteine wie PARP aktiviert und die Apoptose initiiert [83, 179]. Detaillierte Informationen zu den MAPK-, TGF-β- und TNF-α-bezogenen Signalwegen, die Entzündungen, Apoptose und Nekrose induzieren, sind in Abb. 4 zusammengefasst.
Abb. 4
Mitochondriale Schäden
Mitochondrien sind Energieproduktionszentren, die an verschiedenen Signalwegen in Zellen beteiligt sind und auch ein Schlüsselpunkt der apoptotischen Regulation sind [83]. Nach Exposition gegenüber GO und Carboxylgraphen (GXYG) war die Mitochondrienmembran depolarisiert und die Menge an Mitochondrien nahm in HepG2-Zellen ab [180]. Die Exposition gegenüber GFNs führte zu einem signifikant erhöhten gekoppelten und ungekoppelten mitochondrialen Sauerstoffverbrauch, einer Dissipation des mitochondrialen Membranpotentials und schließlich einer Auslösung der Apoptose durch die Aktivierung des mitochondrialen Signalwegs [181]. Beispielsweise erhöhte GO die Aktivität der mitochondrialen Elektronentransportkomplexe I/III und die Zufuhr von Elektronen zu den Stellen I/II der Elektronentransportkette, wodurch die Bildung von ROS während der mitochondrialen Atmung in MHS-Zellen beschleunigt wurde [99]. Die durch GO und das Cytochrom-c/H2O2-Elektronentransfersystem vermittelte Bildung von •OH könnte oxidativen und thermischen Stress verstärken, das mitochondriale Atmungssystem beeinträchtigen und schließlich zu einer dramatischen Toxizität führen [151]. Darüber hinaus könnten die Sauerstoffeinheiten von GO Elektronen von zellulären Redoxproteinen aufnehmen, was den Redoxzyklus von Cytochrom c und Elektronentransportproteinen unterstützt, und die Cytochrome MtrA, MtrB und MtrC/OmcA könnten an der Übertragung von Elektronen auf GO beteiligt sein [182]. Daher können GFNs mit Ausnahme der Plasmamembranschädigung und der Induktion von oxidativem Stress Apoptose und/oder Zellnekrose verursachen, indem sie die mitochondriale Zellaktivität direkt beeinflussen [183, 184].
DNA-Schäden
Aufgrund seiner geringen Größe, großen Oberfläche und Oberflächenladung kann GO signifikante genotoxische Eigenschaften besitzen und schwere DNA-Schäden verursachen, zum Beispiel Chromosomenfragmentierung, DNA-Strangbrüche, Punktmutationen und oxidative DNA-Addukte und -Veränderungen [87, 122, 185 , 186]. Mutagenese wurde bei Mäusen nach intravenöser Injektion von GO in einer Dosis von 20 mg/kg im Vergleich zu Cyclophosphamid (50 mg/kg), einem klassischen Mutagen, beobachtet [112]. Auch wenn GO nicht in den Zellkern eindringen kann, kann es dennoch während der Mitose mit der DNA interagieren, wenn die Kernmembran zerfällt, was die Möglichkeit für DNA-Aberrationen erhöht [87, 147, 187, 188]. Die π-Stacking-Wechselwirkung zwischen den Kohlenstoffringen des Graphens und den hydrophoben DNA-Basenpaaren kann dazu führen, dass ein DNA-Segment auf der Oberfläche von Graphen „aufsteht“ oder „aufliegt“, wobei seine Helixachse senkrecht bzw. parallel ist. Die intermolekularen Kräfte verformen die endständigen Basenpaare der DNA stark, was die Genotoxizität potenziell erhöht [189]. GO kann auch chromosomale Fragmentierung, DNA-Addukte und Punktmutationen induzieren, indem es oxidativen Stress fördert oder Entzündungen durch die Aktivierung intrazellulärer Signalwege wie MAPK, TGF-β und NF-κB auslöst [110, 112, 146]. Graphen und rGO können auch die Expression von p53, Rad51 und MOGG1-1 erhöhen, was Chromosomenschäden widerspiegelt, und die Expression von CDK2 und CDK4 verringern, indem sie den Zellzyklusübergang von der G1- zur S-Phase in verschiedenen Zelllinien stoppen [112 ]. DNA-Schäden können nicht nur die Krebsentstehung initiieren, sondern möglicherweise auch die Gesundheit der nächsten Generation bedrohen, wenn das mutagene Potenzial von GO in Fortpflanzungszellen auftritt, was die Fruchtbarkeit und die Gesundheit der Nachkommen beeinflusst [112, 190].
Entzündungsreaktion
GFNs können in hohen Dosen durch intratracheale Instillation oder intravenöse Verabreichung eine signifikante Entzündungsreaktion einschließlich Entzündungszellinfiltration, Lungenödem und Granulombildung verursachen [30, 49]. Thrombozyten sind die wichtigen Komponenten bei der Gerinnselbildung, um Krankheitserreger und Partikel während der Entzündungsreaktion anzugreifen, und GO könnte direkt die Bildung von plättchenreichen Thromben aktivieren, um Lungengefäße nach intravenöser Injektion zu verschließen [98, 191]. Durch die subkutane Injektion von GO über 21 Tage wurde eine starke Entzündungsreaktion induziert, zusammen mit der Sekretion von Schlüsselzytokinen, einschließlich IL-6, IL-12, TNF-α, MCP-1 und IFN-g [34, 192]. GFNs können eine Entzündungsreaktion und Gewebeschädigung auslösen, indem sie Zytokine und Chemokine freisetzen, die zur Rekrutierung von zirkulierenden Monozyten führen und die Sekretion von Th1/Th2-Zytokinen und Chemokinen stimulieren [124, 193]. Darüber hinaus rufen reines Graphen [193] und rGO [110] eine Entzündungsreaktion hervor, indem sie an Toll-like-Rezeptoren (TLRs) binden und den NF-κB-Signalweg in Zellen aktivieren. Die NF-κB-Signalkaskade wird durch TLRs und proinflammatorische Zytokine wie IL-1 und TNF-α ausgelöst. Bei Aktivierung verschiebt sich NF-κB vom Zytoplasma in den Zellkern, erleichtert die Bindung von abbauendem IκB und wirkt als Transkriptionsfaktor zur Synthese zahlreicher proinflammatorischer Zytokine [194]. Eine schematische Darstellung des Signalweges von TLR4 und TLR9, die durch GFNs aktiviert werden, ist in Abb. 5 gezeigt.
Apoptose
Apoptose ist definiert als die Selbstzerstörung einer Zelle, die von Genen durch komplizierte Programme reguliert wird [83, 195]. GO und rGO verursachten in der Lunge von Mäusen nach Inhalation Apoptose und Entzündung [99], und GFNs hatten auch pro-apoptotische Wirkungen in Zellen [111, 113, 124, 196]. Darüber hinaus schädigten Graphen und GO physikalisch Zellmembranen [166], erhöhten die Permeabilisierung der äußeren mitochondrialen Membran und veränderten das mitochondriale Membranpotential; die erhöhte ROS löste die MAPK- und TGF-β-Signalwege aus und aktivierte Caspase-3 über mitochondrial-abhängige apoptotische Kaskaden, was zur Ausführung der Apoptose führte [83, 99]. In ähnlicher Weise verursachte rGO bei niedriger Dosis und zu einem frühen Zeitpunkt Apoptose, ausgelöst durch den Todesrezeptor und den kanonischen mitochondrialen Signalweg [110]. Eine andere Studie zeigte drei verschiedene Apoptosewege durch GFNs: GO führte durch direkte Interaktion mit Proteinrezeptoren und anschließende Aktivierung des B-Zell-Lymphom-2 (Bcl-2) Wegs zu ROS-abhängiger Apoptose; GO-COOH übertrug ein passives Apoptosesignal an nukleäre DNA, indem es an Proteinrezeptoren bindet und einen ROS-unabhängigen Signalweg aktiviert; GO-PEI schädigte jedoch die Membranen von T-Lymphozyten schwer, um Apoptose auszulösen [105, 197].
Autophagie
Autophagie ist der Prozess des Selbstabbaus von Zellbestandteilen und wurde kürzlich als nicht apoptotischer Zelltod anerkannt [198–200]. Die Autophagie-Aktivierung erfordert die Bildung von Autophagosomen, die Beclin 1, mehrere Autophagie-verwandte Proteine (ATG), Mikrotubuli-assoziierte Protein-Leichtkette 3 (LC3) und p62 enthalten [201]. Die Akkumulation von Autophagosomen ist mit der Exposition gegenüber verschiedenen Nanopartikeln verbunden [202–205], und Autophagie kann extrazelluläre Organismen entfernen und die Organismen im Zytosol zerstören [206]. Es wurde gezeigt, dass GO und GQDs die Akkumulation von Autophagosomen und die Umwandlung von LC3-I in LC3-II induzieren; hemmen den Abbau des autophagischen Substrats p62 Protein [207, 208]. Darüber hinaus kann GO gleichzeitig TLR4- und TLR9-Antworten in Makrophagen [34, 192] und Dickdarmkrebszellen CT26 [206] auslösen. Der Autophagie-Weg ist durch TLR-Signalgebung in Makrophagen mit der Phagozytose verbunden [206, 209].
Nekrose
Nekrose ist eine alternative Form des Zelltods, die durch Entzündungsreaktionen oder Zellverletzungen induziert wird. Die Exposition von Zellen gegenüber reinem Graphen verursacht in hohen Dosen (50 mg/ml) Apoptose und Nekrose [83]. Berichten zufolge führen LDH-Leckage und die Öffnung der mitochondrialen Permeabilitätsübergangspore, die durch einen erhöhten Gehalt an zytoplasmatischem Ca2+ induziert wird, zu Apoptose/Nekrose [210]. Es wurde gezeigt, dass die GO-Behandlung eine makrophagische Nekrose induziert, indem sie die TLR4-Signalgebung aktiviert und anschließend teilweise die autokrine TNF-α-Produktion auslöst [93]. GO in Kombination mit CDDP (GO/CDDP) löste Nekrosen durch Verminderung von RIP1- und Erhöhung von RIP3-Proteinen aus, begleitet von der Freisetzung der Gruppe B1 mit hoher Mobilität (HMGB1) in das Zytosol aus dem Zellkern und aus CT26-Zellen [205, 211, 212].
Epigenetische Veränderungen
Epigenetik umfasst DNA-Methylierung, genomische Prägung, maternale Effekte, Gen-Silencing und RNA-Editierung [213–215]. Die DNA-Methylierung, die eine der am besten untersuchten epigenetischen Modifikationen ist, umfasst Phosphorylierung, Ubiquitinierung und ATP-Ribosylierung und kann zu einem Chromatin-Remodeling führen [197, 216, 217]. Eine kürzlich veröffentlichte Veröffentlichung berichtete, dass die Exposition gegenüber SL-GO/FL-GO zu einer globalen DNA-Hypermethylierung durch Hochregulierung der DNMT3B- und MBD1-Gene führte; Die GNP-Behandlung verursachte eine Hypomethylierung, indem sie die Expression der DNMT3B- und MBD1-Gene verringerte [216]. GO könnte den miRNA-360-Regulationsweg aktivieren, um die DNA-Schadens-Apoptose-Signalkaskade zu unterdrücken, indem es die Komponente von CEP-1 beeinflusst [218]. Zusammengenommen legen diese Daten nahe, dass GFNs durch Modulation epigenetischer Veränderungen subtile Veränderungen in der Genexpressionsprogrammierung verursachen könnten. Es gibt jedoch nur wenige Studien zu GFN-induzierten epigenetischen Veränderungen, und der epigenetische Mechanismus, der durch GFNs-Exposition verursacht wird, ist nicht vollständig verstanden.
Zusammenfassend lässt sich sagen, dass viele Studien repräsentative Mechanismen der GFN-Toxizität diskutiert haben, die vier Signalwege beinhalten: TLRs, TGF-β, TNF-α und MAPKs. Diese vier Signalwege sind korrelativ und kreuzmodulierend, wodurch die Entzündungsreaktion, Autophagie, Apoptose und andere Mechanismen unabhängig und doch miteinander verbunden sind. Darüber hinaus scheint oxidativer Stress die wichtigste Rolle bei der Aktivierung dieser Signalwege zu spielen. Es wurde berichtet, dass es in den Studien zur Toxizität anderer Nanomaterialien Überschneidungen von Apoptose, Autophagie und Nekrose gibt, sie hemmen oder fördern sich unter bestimmten Bedingungen gegenseitig. Die bisher in Veröffentlichungen untersuchten Signalwege der GFN-Toxizität sind jedoch nur ein kleiner Teil eines komplizierten Netzes, und das Netzwerk der Signalwege muss in Zukunft im Detail erforscht werden.
Datenlücken und zukünftige Studien
Derzeit reicht die Literatur nicht aus, um Rückschlüsse auf die potentiellen Gefahren von GFNs zu ziehen. Es haben sich zwei gegensätzliche Meinungen herausgebildet: Einige Forscher schlugen in einer Reihe von Studien mit Schwerpunkt auf biomedizinischen Anwendungen vor, dass Graphenmaterialien biokompatibel sind [119, 154, 162, 219], und andere Studien berichteten über nachteilige biologische Reaktionen und Zytotoxizität [32, 118, 135 , 138, 192]. Diese inkonsistenten Ergebnisse könnten durch mehrere Faktoren verursacht worden sein, darunter die verschiedenen Forschungsgruppen, verschiedene Zell- oder Tiermodelle und unterschiedliche physikalisch-chemische Charakterisierungen von GFNs. Wenn GFNs für in-vivo-Anwendungen im menschlichen Körper oder einige andere biomedizinische Anwendungen erforscht werden, muss die Biokompatibilität berücksichtigt werden, und es sind detailliertere und genauere Studien zur Toxizität von GFNs erforderlich.
Erstens ist eine detaillierte physikalisch-chemische Charakterisierung für alle zukünftigen Studien zur Toxizität von GFNs unerlässlich. In den Experimenten sollten die Merkmalsbeschreibungen von GFNs ihre Größe, Morphologie, Oberfläche, Ladung, Oberflächenmodifikationen, Reinheit und Agglomeration umfassen [88, 141, 148, 162]. Da diese physikalisch-chemischen Faktoren die Toxizität und Biokompatibilität von GFNs stark beeinflussen, sollte ein einfaktorielles Versuchsdesign und der Ausschluss anderer Störfaktoren in Betracht gezogen werden. Details zum Herstellungsprozess sollten ebenfalls angegeben werden, da die gebildeten oxidativen Debris die Oberflächenstruktur von Graphen und GO während der Funktionalisierung stark verändern könnten [151]. Wichtig ist, dass in der Graphentechnologie eine einzige, universelle Methode etabliert werden muss, die einen besseren Vergleich von Daten aus verschiedenen Studien oder verschiedenen Labors ermöglicht.
Zweitens können unterschiedliche Beobachtungskriterien, Parameter und die Auswahl experimenteller Methoden zu großen Variationen zwischen den Labors führen [220, 221]. Beispielsweise kann der MTT-Assay die Graphen-Toxizität immer nicht genau vorhersagen, da die spontane Reduktion zu einem falsch positiven Signal führt. Daher sollten geeignete alternative Bewertungen verwendet werden, wie das wasserlösliche Tetrazoliumsalz-Reagenz (WST-8), der ROS-Assay und der Trypanblau-Ausschlusstest [106, 222]. Darüber hinaus zeigt der Comet-Assay häufig höhere DNA-Schäden als der Mikronukleus-Assay, da ersterer die reparierbare Verletzung und letzterer den nach der Zellteilung verbleibenden Genschaden misst [159, 223]. Daher ist bei der Auswahl des am besten geeigneten Assays zur Bewertung der Toxizität von Graphenmaterialien Vorsicht geboten, um falsch-positive Ergebnisse zu vermeiden.
Drittens ist die Auswahl von Zelllinien von entscheidender Bedeutung, da Krebszelllinien abhängig von ihrem genetischen Hintergrund dazu neigen, empfindlich oder resistent zu sein. Dieselben Graphen-Nanopartikel können je nach ihrer unterschiedlichen Zellherkunft unterschiedliche Reaktionen hervorrufen. Es müssen geeignete Zelllinien mit guter Stabilität verwendet werden, um falsch positive oder negative Ergebnisse zu vermeiden. Von Menschen oder Tieren stammende Primärzellen können den Gesundheitszustand des Menschen besser simulieren. Eine große Menge an Primärzellen wurde verwendet, um die Toxizität anderer Nanomaterialien zu testen [224–228], aber die Kultivierung von Primärzellen ist in den bisherigen Experimenten mit GFNs äußerst selten [210, 229]. Um die physikalisch-chemischen Eigenschaften und die Toxizität von GFNs umfassend zu bewerten, sollten verschiedene Zellexperimente in Kombination mit Primärzellen durchgeführt werden.
Viertens spielt der Verabreichungsweg von GFNs eine sehr wichtige Rolle in Toxizitätsstudien, und unterschiedliche Verabreichungsmethoden führen zu unterschiedlichen toxikologischen Reaktionen [32, 53]. Daher sollten Art und Dauer der Exposition entsprechend dem Ziel der Studie sorgfältig ausgewählt werden. Zur Untersuchung der Neurotoxizität von Nanomaterialien wird häufig die nasale Wirkstoffabgabe verwendet [230, 231], diese Verabreichungsmethode wurde jedoch selten bei der Prüfung der Toxizität von GFNs angewendet. Toxikologische Studien zu GFNs im Nervensystem sind selten, und der Mechanismus ist unklar und muss in Zukunft weiter untersucht werden. Kürzlich durchgeführte toxikokinetische Studien zur Aufnahme, Verteilung, Metabolisierung, Akkumulation und Ausscheidung von GFNs über verschiedene Expositionswege haben einige Ergebnisse geliefert, reichen jedoch bei weitem nicht aus, um die internen komplexen Mechanismen aufzuklären. Zum Beispiel sind weitere Studien erforderlich, um die spezifischen molekularen Mechanismen von GFNs, die die physiologischen Barrieren passieren, und die Menge der Akkumulation oder die Ausscheidungszeit von GFNs im Gewebe zu verstehen. Darüber hinaus ist angesichts der erhöhten Exposition des Menschen gegenüber GFN die Bewertung der systemischen Toxizität im menschlichen Körper für zukünftige Studien unverzichtbar.
Fünftens ist ein weiteres wichtiges Thema, das Aufmerksamkeit erfordert, das langfristige Schicksal von GFNs, nachdem sie in den Körper gelangt oder von Zellen aufgenommen wurden. Die jüngsten Studien bestanden aus Bewertungen der Kurzzeittoxizität [89, 232], und toxische Langzeitschäden haben seit der weit verbreiteten Anwendung von GFNs im Jahr 2008 nicht viel Beachtung gefunden. Darüber hinaus kann eine funktionalisierte Graphenoberfläche ihre Biokompatibilität verbessern, aber die
Sechstens müssen spezifischere Signalwege im Mechanismus der GFN-Toxizität entdeckt und aufgeklärt werden. Derzeit wurden mehrere typische Toxizitätsmechanismen von GFNs veranschaulicht und allgemein akzeptiert, wie z. B. oxidativer Stress, Apoptose und Autophagie. Diese Mechanismen wurden jedoch nur allgemein beschrieben und die spezifischen Signalwege innerhalb dieser Mechanismen müssen im Detail untersucht werden. Auch die Signalwege, die an der Toxizität anderer Nanomaterialien beteiligt sind, können für die Untersuchung von GFNs von Bedeutung sein. Daher sollten in der zukünftigen Forschung weitere Signalwege entdeckt werden. So wurde beispielsweise die Nanoepigenetik in zahlreichen Studien zu Nanomaterialien berücksichtigt, was auch bei der Beurteilung der begrenzten Toxizität und Nebenwirkungen von GFNs hilfreich ist. Jüngste Studien haben gezeigt, dass GFNs epigenetische und genomische Veränderungen verursachen können, die physikalische Toxizität und Karzinogenität stimulieren können [234]. GFNs haben große Oberflächen, glatte, kontinuierliche Oberflächen und Bio-Persistenz, ähnlich den Eigenschaften von tumorigenen Festkörperimplantaten. Es ist nicht bekannt, ob GFNs das Potenzial haben, Fremdkörpersarkome zu induzieren, und daher sollten so bald wie möglich definitive Studien zu Tumorpotenzialen oder Risiken von Graphen durchgeführt werden.
Schlussfolgerungen
In den letzten Jahren wurden GFNs in zahlreichen technologischen und biomedizinischen Bereichen eingesetzt. Derzeit haben sich die meisten Experimente auf die Toxizität von GFNs in Lunge und Leber konzentriert. Daher verdienen Studien zu Hirnverletzungen oder Neurotoxizität in Zukunft mehr Aufmerksamkeit. Viele Experimente haben gezeigt, dass GFNs bei vielen biologischen Anwendungen toxische Nebenwirkungen haben, aber die eingehende Untersuchung der Toxizitätsmechanismen ist dringend erforderlich. Darüber hinaus müssen gegensätzliche Ergebnisse zur Toxizität von GFNs durch wirksame experimentelle Methoden und systematische Studien angegangen werden. Dieser Review bietet einen Überblick über die Toxizität von GFNs, indem er die Toxikokinetik, Toxizitätsmechanismen und Einflussfaktoren zusammenfasst und Informationen liefern soll, um in Zukunft eine gründliche Erforschung der Hämo- und Biokompatibilität von GFNs in vitro und in vivo zu erleichtern. Diese Überprüfung wird dazu beitragen, Sicherheitsbedenken vor der klinischen und therapeutischen Anwendung von GFNs auszuräumen, was für die weitere Entwicklung von GFNs in biologischen Anwendungen wichtig sein wird.
Abkürzungen
AM:
Alveoläre Makrophagen
BBB:
Blut-Hirn-Schranke
BEB:
Blut-Nebenhoden-Schranken
BTB:
Blut-Hoden-Schranke
CR:
Komplementrezeptor
FcgR:
Fcg-Rezeptor
BLG:
Wenigschichtiges Graphen
GFNs:
Nanomaterialien der Graphen-Familie
GNS:
Graphen-Nanoblätter
GEHEN:
Graphenoxid
GO-COOH:
Carboxyliertes Graphenoxid
GO-DEX:
GO-Dextran
GO-MB:
GO-molekulares Leuchtfeuer
GO-NH2:
Aminiertes GO
GO-PAA:
Poly(acrylsäure)-funktionalisiertes GO
GO-PAM:
Poly(acrylamid)-funktionalisiertes GO
GO-PEG:
PEGylierte GO-Derivate
GO-PEI:
GO-Polyethylenimin
GQDs:
Graphen-Quantenpunkte
GSH-PX:
Glutathionperoxidase
GXVG:
Carboxylgraphen
LDH:
Laktat und Dehydrogenase
MALDI:
Matrixunterstützte Laserdesorption/Ionisation
MAPKs:
Mitogen-aktivierte Proteinkinase
MDA:
Malondialdehyd
MØ:
Makrophage
HERR:
Mannose-Rezeptor
MSI:
Massenspektrometrie-Bildgebung
PC12-Zellen:
Phäochromozytomzellen der Ratte
PCGO:
Proteinbeschichtete Graphenoxid-Nanopartikel
PrGO:
PEGyliertes reduziertes Graphenoxid
RES:
Retikuloendotheliales System
rGO:
Reduziertes Graphenoxid
ROS:
Reaktive Sauerstoffspezies
SOD:
Hyperventilieren
TLRs:
Toll-like-Rezeptor
Verweise
1.
KS Novoselov, AK Geim, SV Morozov, D. Jiang, Y. Zhang, SV Dubonos et al. Elektrischer Feldeffekt in atomar dünnen Kohlenstoffschichten. Wissenschaft. 2004;306(5696):666–9.
CAS
PubMed
Artikel
Google Scholar
2.
Sanchez VC, Jachak A, Hurt RH, Kane AB. Biologische Wechselwirkungen von Nanomaterialien der Graphenfamilie: eine interdisziplinäre Übersicht. Chem. Res. Toxicol. 2012;25(1):15–34.
CAS
PubMed
Artikel
Google Scholar
3.
Yang XY, Wang YS, Huang X, Ma YF, Huang Y, Yang RC, et al. Multifunktionalisierter Antikrebs-Wirkstoffträger auf Graphenoxidbasis mit Dual-Targeting-Funktion und pH-Sensitivität. J Mat Chem. 2011;21(10):3448–54.
CAS
Artikel
Google Scholar
4.
Park S, An J, Jung I, Piner RD, An SJ, Li X, et al. Kolloidale Suspensionen von stark reduziertem Graphenoxid in einer Vielzahl von organischen Lösungsmitteln. Nano Lett. 2009;9(4):1593–7.
CAS
PubMed
Artikel
Google Scholar
5.
Geim AK. Graphen: Status und Perspektiven. Wissenschaft. 2009;324(5934)::1530–4.
CAS
PubMed
Artikel
Google Scholar
6.
Guo X, Mei N. Bewertung des toxischen Potenzials von Nanomaterialien der Graphenfamilie. J Nahrungsmittelarzneimittel-Anal. 2014;22(1):105–15.
CAS
PubMed
Artikel
Google Scholar
7.
Seabra AB, Paula AJ, de Lima R, Alves OL, Duran N. Nanotoxizität von Graphen und Graphenoxid. Chem. Res. Toxicol. 2014;27(2):159–68.
CAS
PubMed
Artikel
Google Scholar
8.
Shen H, Zhang L, Liu M, Zhang Z. Biomedizinische Anwendungen von Graphen. Theranostik. 2012;2(3):283-94.
CAS
PubMed
PubMed-Zentrale
Artikel
Google Scholar
9.
Han
Ms:
-
Alveolar macrophages
- BBB:
-
Blood-brain barrier
- BEB:
-
Blood-epididymis barriers
- BTB:
-
Blood-testis barrier
- CR:
-
Complement receptor
- FcgR:
-
Fcg receptor
- FLG:
-
Few-layer graphene
- GFNs:
-
Graphene family nanomaterials
- GNS:
-
Graphene nanosheets
- GO:
-
Graphene oxide
- GO-COOH:
-
Carboxylated graphene oxide
- GO-DEX:
-
GO-dextran
- GO-MB:
-
GO-molecular beacon
- GO-NH2:
-
Aminated GO
- GO-PAA:
-
Poly(acrylic acid)-functionalized GO
- GO-PAM:
-
Poly(acrylamide)-functionalized GO
- GO-PEG:
-
PEGylated GO derivatives
- GO-PEI:
-
GO-polyethylenimine
- GQDs:
-
Graphene quantum dots
- GSH-PX:
-
Glutathione peroxidase
- GXVG:
-
Carboxyl graphene
- LDH:
-
Lactate and dehydrogenase
- MALDI:
-
Matrix-assisted laser desorption/ionization
- MAPKs:
-
Mitogen-activated protein kinase
- MDA:
-
Malondialdehyde
- MØ:
-
Macrophage
- MR:
-
Mannose receptor
- MSI:
-
Mass spectrometry imaging
- PC12 cells:
-
Rat pheochromocytoma cells
- PCGO:
-
Protein-coated graphene oxide nanoparticles
- PrGO:
-
PEGylated reduced graphene oxide
- RES:
-
Reticuloendothelial system
- rGO:
-
Reduced graphene oxide
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
- TLRs:
-
Toll-like receptor
References
- 1.
Novoselov KS, Geim AK, Morozov SV, Jiang D, Zhang Y, Dubonos SV, et al. Electric field effect in atomically thin carbon films. Science. 2004;306(5696):666–9.
- 2.
Sanchez VC, Jachak A, Hurt RH, Kane AB. Biological interactions of graphene-family nanomaterials: an interdisciplinary review. Chem Res Toxicol. 2012;25(1):15–34.
- 3.
Yang XY, Wang YS, Huang X, Ma YF, Huang Y, Yang RC, et al. Multi-functionalized graphene oxide based anticancer drug-carrier with dual-targeting function and pH-sensitivity. J Mat Chem. 2011;21(10):3448–54.
- 4.
Park S, An J, Jung I, Piner RD, An SJ, Li X, et al. Colloidal suspensions of highly reduced graphene oxide in a wide variety of organic solvents. Nano Lett. 2009;9(4):1593–7.
- 5.
Geim AK. Graphene: status and prospects. Science. 2009;324(5934):1530–4.
- 6.
Guo X, Mei N. Assessment of the toxic potential of graphene family nanomaterials. J Food Drug Anal. 2014;22(1):105–15.
- 7.
Seabra AB, Paula AJ, de Lima R, Alves OL, Duran N. Nanotoxicity of graphene and graphene oxide. Chem Res Toxicol. 2014;27(2):159–68.
- 8.
Shen H, Zhang L, Liu M, Zhang Z. Biomedical applications of graphene. Theranostics. 2012;2(3):283–94.
- 9.
Han U, Seo Y, Hong J. Effect of pH on the structure and drug release profiles of layer-by-layer assembled films containing polyelectrolyte, micelles, and graphene oxide. Sci Rep. 2016;6(2045–2322 (Electronic)):24158.
- 10.
Wang H, Liang Y, Mirfakhrai T, Chen Z, Casalongue HS, Dai H. Advanced asymmetrical supercapacitors based on graphene hybrid materials. Nano Res. 2011;4(8):729–36.
- 11.
Loh KP, Bao Q, Eda G, Chhowalla M. Graphene oxide as a chemically tunable platform for optical applications. Nat Chem. 2010;2(12):1015–24.
- 12.
Wang D, Zhu L, Chen JF, Dai L. Mn3O4-graphene hybrid as a high-capacity anode material for lithium ion batteries. J Am Chem Soc. 2015;132(1520–5126 (Electronic)):13978–80.
- 13.
Gurunathan S, Han JW, Dayem AA, Eppakayala V, Kim JH. Oxidative stress-mediated antibacterial activity of graphene oxide and reduced graphene oxide in Pseudomonas aeruginosa. Int J Nanomed. 2012;7(1178–2013 (Electronic)):e14.
- 14.
Zhan S, Zhu D, Ma S, Yu W, Jia Y, Li Y, et al. Highly efficient removal of pathogenic bacteria with magnetic graphene composite. ACS Appl Mater Interf. 2015;7(1944–8252 (Electronic)):4290–8.
- 15.
Yang HW, Hua MY, Chen SL, Tsai RY. Reusable sensor based on high magnetization carboxyl-modified graphene oxide with intrinsic hydrogen peroxide catalytic activity for hydrogen peroxide and glucose detection. Biosens Bioelectron. 2013;41:172–9.
- 16.
Wang Y, Yuan R, Chai Y, Yuan Y, Bai L. In situ enzymatic silver enhancement based on functionalized graphene oxide and layer-by-layer assembled gold nanoparticles for ultrasensitive detection of thrombin. Biosens Bioelectron. 2012;38(1):50–4.
- 17.
Huang J, Zhang L, Liang RP, Qiu JD. “On-off” switchable electrochemical affinity nanobiosensor based on graphene oxide for ultrasensitive glucose sensing. Biosens Bioelectron. 2013;41:430–5.
- 18.
Gao L, Lian C, Zhou Y, Yan L, Li Q, Zhang C, et al. Graphene oxide-DNA based sensors. Biosens Bioelectron. 2014;60(1873–4235 (Electronic)):22–9.
- 19.
Chen ML, Liu JW, Hu B, Chen ML, Wang JH. Conjugation of quantum dots with graphene for fluorescence imaging of live cells. Analyst. 2011;136(20):4277–83.
- 20.
Wang Y, Wang H, Liu D, Song S, Wang X, Zhang H. Graphene oxide covalently grafted upconversion nanoparticles for combined NIR mediated imaging and photothermal/photodynamic cancer therapy. Biomaterials. 2013;34(1878–5905 (Electronic)):7715–24.
- 21.
Pan Y, Sahoo NG, Li L. The application of graphene oxide in drug delivery. Expert Opin Drug Deliv. 2012;9(11):1365–76.
- 22.
Huiyun W, Chunyan D, Haiqing D, Aijun S, Wenjuan X, Xiaojun C, et al. Engineered redox-responsive PEG detachment mechanism in PEGylated nano-graphene oxide for intracellular drug delivery. Small. 2012;8(5):760–9.
- 23.
Yang X, Qiu L, Cheng C, Wu Y, Ma ZF, Li D. Ordered gelation of chemically converted graphene for next-generation electroconductive hydrogel films. Angewandte Chem Int Ed Engl. 2011;50(32):7325–8.
- 24.
Schinwald A, Murphy FA, Jones A, Macnee W, Donaldson K. Graphene-based nanoplatelets: a new risk to the respiratory system as a consequence of their unusual aerodynamic properties. ACS Nano. 2012;6(1):736–46.
- 25.
Chaenyung C, Ryon SS, Xiguang G, Nasim A, Dokmeci MR, Xiaowu Shirley T, et al. Controlling mechanical properties of cell-laden hydrogels by covalent incorporation of graphene oxide. Small. 2014;10(3):514–23.
- 26.
Arvidsson R, Molander S, Sandén BA. Review of potential environmental and health risks of the nanomaterial graphene. Hum Ecol Risk Assess. 2013;19(4):873–87.
- 27.
Lee JH, Han JH, Kim JH, Kim B, Bello D, Kim JK, et al. Exposure monitoring of graphene nanoplatelets manufacturing workplaces. Inhal Toxicol. 2016;28(6):281–91.
- 28.
Maynard RL. Nano-technology and nano-toxicology. Emerg Health Threats J. 2012;5.
- 29.
Su WC, Ku BK, Kulkarni P, Cheng YS. Deposition of graphene nanomaterial aerosols in human upper airways. J Occup Environ Hyg. 2015;13(1):1–34.
- 30.
Li B, Yang J, Huang Q, Zhang Y, Peng C, Zhang Y, et al. Biodistribution and pulmonary toxicity of intratracheally instilled graphene oxide in mice. NPG Asia Mater. 2013;5:E44.
- 31.
Yang K, Gong H, Shi X, Wan J, Zhang Y, Liu Z. In vivo biodistribution and toxicology of functionalized nano-graphene oxide in mice after oral and intraperitoneal administration. Biomaterials. 2013;34(11):2787–95.
- 32.
Wen KP, Chen YC, Chuang CH, Chang HY, Lee CY, Tai NH. Accumulation and toxicity of intravenously-injected functionalized graphene oxide in mice. J Appl Toxicol. 2015;35(10):1211–8.
- 33.
Kurantowicz N, Strojny B, Sawosz E, Jaworski S, Kutwin M, Grodzik M, et al. Biodistribution of a high dose of diamond, graphite, and graphene oxide nanoparticles after multiple intraperitoneal injections in rats. Nanoscale Res Lett. 2015;10(1):398.
- 34.
Yue H, Wei W, Yue Z, Wang B, Luo N, Gao Y, et al. The role of the lateral dimension of graphene oxide in the regulation of cellular responses. Biomaterials. 2012;33(16):4013–21.
- 35.
Nezakati T, Cousins BG, Seifalian AM. Toxicology of chemically modified graphene-based materials for medical application. Arch Toxicol. 2014;88(11):1987–2012.
- 36.
Chng ELK, Pumera M. Toxicity of graphene related materials and transition metal dichalcogenides. Rsc Advances. 2015;5(4):3074–80.
- 37.
Zheng XT, Ananthanarayanan A, Luo KQ, Chen P. Glowing graphene quantum dots and carbon dots: properties, syntheses, and biological applications. Small. 2015;11(1613–6829 (Electronic)):1620–36.
- 38.
Caffo M, Merlo L, Marino D, Caruso G. Graphene in neurosurgery: the beginning of a new era. Nanomed. 2015;10:615–25.
- 39.
Wu SY, An SS, Hulme J. Current applications of graphene oxide in nanomedicine. Int J Nanomed. 2015;10(Spec Iss):9–24.
- 40.
Tonelli FMP, Goulart VAM, Gomes KN, Ladeira MS, Santos AK, Lorencon E, et al. Graphene-based nanomaterials: biological and medical applications and toxicity. Nanomedicine. 2015;10(15):2423–50.
- 41.
Zhou R, Gao H. Cytotoxicity of graphene: recent advances and future perspective. Wiley Interdiscip Rev Nanomed Nanobiotechnol. 2014;6(5):452–74.
- 42.
Ema M, Hougaard KS, Kishimoto A, Honda K. Reproductive and developmental toxicity of carbon-based nanomaterials: A literature review. Nanotoxicology. 2015;10:391–412.
- 43.
Jastrzebska AM, Olszyna AR. The ecotoxicity of graphene family materials: current status, knowledge gaps and future needs. J Nanopart Res. 2015;17(1):1–21.
- 44.
Xu S, Zhang Z, Chu M. Long-term toxicity of reduced graphene oxide nanosheets: Effects on female mouse reproductive ability and offspring development. Biomaterials. 2015;54:188–200.
- 45.
Jennifer M, Maciej W. Nanoparticle technology as a double-edged sword: cytotoxic, genotoxic and epigenetic effects on living cells. J Biomater Nanobiotechnol. 2013;4:53–63.
- 46.
Wu W, Yan L, Wu Q, Li Y, Li Q, Chen S, et al. Evaluation of the toxicity of graphene oxide exposure to the eye. Nanotoxicology. 2016;10(9):1329–40.
- 47.
Lee K, Jeong Y, Bae J, Seok H, Yang Y, Roh S, et al. The role of surface functionalization on the pulmonary inflammogenicity and translocation into mediastinal lymph nodes of graphene nanoplatelets in rats. Arch Toxicol.2016:1–10.
- 48.
Schinwald A, Murphy F, Askounis A, Koutsos V, Sefiane K, Donaldson K, et al. Minimal oxidation and inflammogenicity of pristine graphene with residence in the lung. Nanotoxicology. 2013;8(8):824–32.
- 49.
Zhang X, Yin J, Peng C, Hu W, Zhu Z, Li W, et al. Distribution and biocompatibility studies of graphene oxide in mice after intravenous administration. Carbon. 2011;49(3):986–95.
- 50.
Singh SK, Singh MK, Nayak MK, Kumari S, Shrivastava S, Gracio JJ, et al. Thrombus inducing property of atomically thin graphene oxide sheets. ACS Nano. 2011;5(6):4987–96.
- 51.
Gurunathan S, Han JW, Eppakayala V, Kim JH. Biocompatibility of microbially reduced graphene oxide in primary mouse embryonic fibroblast cells. Colloids Surf B Biointerf. 2013;105:58–66.
- 52.
Yang K, Wan J, Zhang S, Zhang Y, Lee ST, Liu Z. In vivo pharmacokinetics, long-term biodistribution, and toxicology of PEGylated graphene in mice. ACS Nano. 2011;5(1):516–22.
- 53.
Fu C, Liu T, Li L, Liu H, Liang Q, Meng X. Effects of graphene oxide on the development of offspring mice in lactation period. Biomaterials. 2015;40:23–31.
- 54.
Hu Q, Jiao B, Shi X, Valle RP, Zuo YY, Hu G. Effects of graphene oxide nanosheets on the ultrastructure and biophysical properties of the pulmonary surfactant film. Nanoscale. 2015;7(43):18025–9.
- 55.
Gosens I, Post JA, de la Fonteyne LJ, Jansen EH, Geus JW, Cassee FR, et al. Impact of agglomeration state of nano- and submicron sized gold particles on pulmonary inflammation. Part Fibre Toxicol. 2010;7(1743–8977 (Electronic)):1.
- 56.
Geiser M, Kreyling WG. Deposition and biokinetics of inhaled nanoparticles. Part Fibre Toxicol. 2010;7:2.
- 57.
Ruge CA, Schaefer UF, Herrmann J, Kirch J, Canadas O, Echaide M, et al. The interplay of lung surfactant proteins and lipids assimilates the macrophage clearance of nanoparticles. PLoS One. 2012;7(7):e40775.
- 58.
Morfeld P, Treumann S, Ma-Hock L, Bruch J, Landsiedel R. Deposition behavior of inhaled nanostructured TiO2 in rats: fractions of particle diameter below 100 nm (nanoscale) and the slicing bias of transmission electron microscopy. Inhal Toxicol. 2012;24(1091–7691 (Electronic)):939–51.
- 59.
Wiemann M, Vennemann A, Sauer UG, Wiench K, Ma-Hock L, Landsiedel R. An in vitro alveolar macrophage assay for predicting the short-term inhalation toxicity of nanomaterials. J Nanobiotechnol. 2016;14(1477–3155 (Electronic)):1.
- 60.
Kreyling WG, Semmler-Behnke M, Takenaka S, Möller W. Differences in the biokinetics of inhaled nano- versus micrometer-sized particles. Accounts Chem Res. 2012;46(1520–4898 (Electronic)):714–22.
- 61.
Liang M, Hu M, Pan B, Xie Y, Petersen EJ. Biodistribution and toxicity of radio-labeled few layer graphene in mice after intratracheal instillation. Part Fibre Toxicol. 2016;13(1):1–12.
- 62.
Abbott NJ, Patabendige AA, Dolman DE, Yusof SR, Begley DJ. Structure and function of the blood–brain barrier. Neurobiol Dis. 2010;37(1):13–25.
- 63.
Mendonca MC, Soares ES, de Jesus MB, Ceragioli HJ, Ferreira MS, Catharino RR, et al. Reduced graphene oxide induces transient blood–brain barrier opening: an in vivo study. J Nanobiotechnol. 2015;13:78.
- 64.
Liu Y, Xu LP, Dai W, Dong H, Wen Y, Zhang X. Graphene quantum dots for the inhibition of beta amyloid aggregation. Nanoscale. 2015;7(45):19060–5.
- 65.
Mital P, Hinton BT, Dufour JM. The blood-testis and blood-epididymis barriers are more than just their tight junctions. Biol Reprod. 2011;84(5):851–8.
- 66.
Liang S, Xu S, Zhang D, He J, Chu M. Reproductive toxicity of nanoscale graphene oxide in male mice. Nanotoxicology. 2015;9(1):92–105.
- 67.
Buerkithurnherr T, Von MU, Wick P. Knocking at the door of the unborn child: engineered nanoparticles at the human placental barrier. Swiss Med Wkly. 2012;142:w13559.
- 68.
Yang H, Sun C, Fan Z, Tian X, Yan L, Du L, et al. Effects of gestational age and surface modification on materno-fetal transfer of nanoparticles in murine pregnancy. Sci Rep. 2012;2(46):847.
- 69.
Huang X, Zhang F, Sun X, Choi KY, Niu G, Zhang G, et al. The genotype-dependent influence of functionalized multiwalled carbon nanotubes on fetal development. Biomaterials. 2014;35(2):856–65.
- 70.
Qi W, Bi J, Zhang X, Wang J, Wang J, Liu P, et al. Damaging effects of multi-walled carbon nanotubes on pregnant mice with different pregnancy times. Sci Rep. 2014;4(3):doi: 10.1038/srep04352.
- 71.
Du J, Wang S, You H, Jiang R, Zhuang C, Zhang X. Developmental toxicity and DNA damage to zebrafish induced by perfluorooctane sulfonate in the presence of ZnO nanoparticles. Environ Toxicol. 2014;31(1522–7278 (Electronic)):360–71.
- 72.
Zhou Z, Son J, Harper B, Zhou Z, Harper S. Influence of surface chemical properties on the toxicity of engineered zinc oxide nanoparticles to embryonic zebrafish. Beilstein J Nanotechnol. 2015;6(2190–4286 (Electronic)):1568–79.
- 73.
Rollerova E, Tulinska J, Liskova A, Kuricova M, Kovriznych J, Mlynarcikova A, et al. Titanium dioxide nanoparticles: some aspects of toxicity/focus on the development. Endocr Reg. 2014;49(1210–0668 (Print)):97–112.
- 74.
Warheit DB, Boatman R, Brown SC. Developmental toxicity studies with 6 forms of titanium dioxide test materials (3 pigment-different grade & 3 nanoscale) demonstrate an absence of effects in orally-exposed rats. Reg Toxicol Pharmacol. 2015;73(1096–0295 (Electronic)):887–96.
- 75.
Ema M, Gamo M, Honda K. Developmental toxicity of engineered nanomaterials in rodents. Toxicol Appl Pharmacol. 2015;299(1096–0333 (Electronic)):47–52.
- 76.
Li Z, Geng Y, Zhang X, Qi W, Fan Q, Li Y, et al. Biodistribution of co-exposure to multi-walled carbon nanotubes and graphene oxide nanoplatelets radiotracers. J Nanopart Res. 2011;13(7):2939–47.
- 77.
Wang Y, Li Z, Hu D, Lin CT, Li J, Lin Y. Aptamer/graphene oxide nanocomplex for in situ molecular probing in living cells. J Am Chem Soc. 2010;132(27):9274–6.
- 78.
Liu JH, Yang ST, Wang H, Chang Y, Cao A, Liu Y. Effect of size and dose on the biodistribution of graphene oxide in mice. Nanomedicine. 2012;7(12):1801–12.
- 79.
Zhang S, Yang K, Feng L, Liu Z. In vitro and in vivo behaviors of dextran functionalized graphene. Carbon. 2011;49(12):4040–9.
- 80.
Hirn S, Semmler-Behnke M, Schleh C, Wenk A, Lipka J, Schaffler M, et al. Particle size-dependent and surface charge-dependent biodistribution of gold nanoparticles after intravenous administration. Eur J Pharm Biopharm. 2011;77(3):407–16.
- 81.
Li B, Zhang XY, Yang JZ, Zhang YJ, Li WX, Fan CH, et al. Influence of polyethylene glycol coating on biodistribution and toxicity of nanoscale graphene oxide in mice after intravenous injection. Int J Nanomedicine. 2014;9:4697–707.
- 82.
Zhang Y, Ali SF, Dervishi E, Xu Y, Li Z, Casciano D, et al. Cytotoxicity effects of graphene and single-wall carbon nanotubes in neural phaeochromocytoma-derived PC12 cells. ACS Nano. 2010;4(6):3181–6.
- 83.
Li Y, Liu Y, Fu Y, Wei T, Le Guyader L, Gao G, et al. The triggering of apoptosis in macrophages by pristine graphene through the MAPK and TGF-beta signaling pathways. Biomaterials. 2012;33(2):402–11.
- 84.
Sydlik SA, Jhunjhunwala S, Webber MJ, Anderson DG, Langer R. In vivo compatibility of graphene oxide with differing oxidation states. ACS Nano. 2015;9(4):3866–74.
- 85.
Mytych J, Wnuk M. Nanoparticle technology as a double-edged sword: cytotoxic, genotoxic and epigenetic effects on living cells. J Biomater Nanobiotechnol. 2013;4:53–63.
- 86.
Peng C, Hu W, Zhou Y, Fan C, Huang Q. Intracellular imaging with a graphene-based fluorescent probe. Small. 2010;6(15):1686–92.
- 87.
Wang D, Zhu L, Chen JF, Dai L. Can graphene quantum dots cause DNA damage in cells? Nanoscale. 2015;7(21):9894–901.
- 88.
Mu Q, Su G, Li L, Gilbertson BO, Yu LH, Zhang Q, et al. Size-dependent cell uptake of protein-coated graphene oxide nanosheets. ACS Appl Mater Interf. 2012;4(4):2259–66.
- 89.
Xu M, Zhu J, Wang F, Xiong Y, Wu Y, Wang Q, et al. Improved in vitro and in vivo biocompatibility of graphene oxide through surface modification: poly(acrylic acid)-functionalization is superior to PEGylation. ACS Nano. 2016;10:3267–81.
- 90.
Kostarelos K, Novoselov KS. Materials science. Exploring the interface of graphene and biology. Science. 2014;344(6181):261–3.
- 91.
Sasidharan A, Panchakarla LS, Chandran P, Menon D, Nair S, Rao CN, et al. Differential nano-bio interactions and toxicity effects of pristine versus functionalized graphene. Nanoscale. 2011;3(6):2461–4.
- 92.
Li Y, Yuan H, von dem Bussche A, Creighton M, Hurt RH, Kane AB, et al. Graphene microsheets enter cells through spontaneous membrane penetration at edge asperities and corner site. Proc Natl Acad Sci U S A. 2013;110(1091–6490 (Electronic)):12295–300.
- 93.
Qu G, Liu S, Zhang S, Wang L, Wang X, Sun B, et al. Graphene oxide induces toll-like receptor 4 (TLR4)-dependent necrosis in macrophages. ACS Nano. 2013;7(7):5732–45.
- 94.
Ma J, Liu R, Wang X, Liu Q, Chen Y, Valle RP, et al. Crucial role of lateral size for graphene oxide in activating macrophages and stimulating Pro-inflammatory responses in cells and animals. ACS Nano. 2015;9(10):10498–515.
- 95.
Mao L, Hu M, Pan B, Xie Y, Petersen EJ. Biodistribution and toxicity of radio-labeled few layer graphene in mice after intratracheal instillation. Part Fibre Toxicol. 2016;13(1743–8977 (Electronic)):1.
- 96.
Park EJ, Lee SJ, Lee K, Choi YC, Lee BS, Lee GH, et al. Pulmonary persistence of graphene nanoplatelets may disturb physiological and immunological homeostasis. J Appl Toxicol. 2016.
- 97.
Kim JK, Shin JH, Lee JS, Hwang JH, Lee JH, Baek JE, et al. 28-Day inhalation toxicity of graphene nanoplatelets in Sprague–Dawley rats. Nanotoxicology. 2016;10(7):891–901.
- 98.
Singh SK, Singh MK, Kulkarni PP, Sonkar VK, Gracio JJ, Dash D. Amine-modified graphene: thrombo-protective safer alternative to graphene oxide for biomedical applications. ACS Nano. 2012;6(3):2731–40.
- 99.
Duch MC, Budinger GR, Liang YT, Soberanes S, Urich D, Chiarella SE, et al. Minimizing oxidation and stable nanoscale dispersion improves the biocompatibility of graphene in the lung. Nano Lett. 2011;11(12):5201–7.
- 100.
Wang X, Duch MC, Mansukhani N, Ji Z, Liao YP, Wang M, et al. Use of a pro-fibrogenic mechanism-based predictive toxicological approach for tiered testing and decision analysis of carbonaceous nanomaterials. ACS Nano. 2015;9(1936-086X (Electronic)):3032–43.
- 101.
Sawosz E, Jaworski S, Kutwin M, Hotowy A, Wierzbicki M, Grodzik M, et al. Toxicity of pristine graphene in experiments in a chicken embryo model. Int J Nanomed. 2014;9:3913–22.
- Weiter im Link https://particleandfibretoxicology.biomedcentral.com/articles/10.1186/s12989-016-0168-y#Fig1https://particleandfibretoxicology.biomedcentral.com/articles/10.1186/s12989-016-0168-y#Fig1
Seite 302 von 1330